Близнецы [Иван Иванович Канаев] (fb2) читать онлайн
- Близнецы (а.с. 1) 77.34 Мб, 339с. скачать: (fb2) читать: (полностью) - (постранично) - Иван Иванович Канаев
[Настройки текста] [Cбросить фильтры]
[Оглавление]
ПРЕДИСЛОВИЕ
Настоящая книга является первой попыткой составить на русском языке краткий обзор огромного материала по изучению близнецов. Литература о близнецах насчитывает несколько тысяч названий и находится преимущественно в различных журналах порой трудно доступных. Новейшей, самой объемистой сводкой по близнецам является книга Гедды (Джедда — Gedda, 1951), имеющая 1381 страницу, из которых 239 страниц содержат список литературы. Дополнение и продолжение этого списка дает Фершюр (Verschuer) в своей книге 1954 г. В обоих списках названо более семи тысяч работ по близнецам, причем эти списки не являются исчерпывающими. Кроме того, на иностранных языках существует несколько сравнительно кратких обзорных работ, служащих своего рода введением в науку о близнецах, например книги Лотце (Lotze, 1937), Ньюмена (Newman, 1940), Коллери (Gaullery, 1945) и другие. Вопросы, которые затрагиваются изучением близнецов, факты, которые получены в этой области, «близнецовый метод» генетики — все это должно быть использовано специалистами в разных областях биологии и может стимулировать решение ряда очередных задач советской науки. Книга расчитана на читателей разных специальностей: зоологов, эмбриологов, генетиков, антропологов, этнографов, физиологов, психологов, педагогов, врачей. Проблема близнечества не лишена интереса и для историков культуры, искусствоведов, литературоведов, а также для широкого круга читателей.И. Канаев. Ленинград. Март 1957 г.
ВВЕДЕНИЕ
Наука о близнецах, которую недавно некоторые ученые стали называть «гемеллологией», за последние десятилетия нашего века очень интенсивно развивалась, особенно в области генетики, эмбриологии и медицины. Близнецы как объект по-разному изучались различными дисциплинами в соответствии с их задачами и методами; но вместе с тем каждая из них внесла свой вклад в понимание близнецов как явления природы, и познание проблемы близнечества. Близнецами обычно называют потомство одной матери, развившееся совместно в одной утробе и потому родившееся более или менее одновременно: у животных, детенышей одного помета, у человека — детей, появившихся на свет за одни роды. Иначе говоря, под словом «близнецы» чаще всего подразумевают потомство живородящих млекопитающих, зачастую в норме одноплодных, как слон, корова, лошадь, человек. По числу родившихся близнецов можно говорить о двойнях, когда близнецов два, о тройнях, когда их три, и т. д. Нередко слово «близнецы» употребляется как синоним двойни, так как близнецы чаще всего бывают именно двойнями. В некоторых языках в самом слове, которым обозначаются близнецы, содержится корень слова «два», как в русском слове «двойня»; например, в немецком слове «Zwillinge» (корень «zwei»— два) или английском слове «twins» (корень «two» — два). Поэтому логично для английского языка называть троен и других близнецов, более многочисленных, термином «сверхблизнецы» («super-twins»), как это предложил Ньюмен; для русского же языка это не имеет смысла. В науке, однако, термин «близнецы» употребляется в гораздо более широком смысле, чем в обычной речи. Близнецы описаны не только у различных групп низших, беспозвоночных животных, но также и у растений. Это объясняется существенными чертами сходства в способах возникновения близнецов у разных групп организмов, филогенетически порой очень далеко стоящих друг от друга. По способу возникновения различаются два основных типа близнецов: однояйцевые близнецы (ОБ) и разнояйцевые близнецы (РБ), или двуяйцевые. Первые возникают из одного яйца, оплодотворенного одним спермием; это, в сущности, результат бесполого размножения оплодотворенного яйца (зиготы) в период дробления или зародыша на ранних стадиях развития. Иначе говоря, из одного оплодотворенного яйца вместо полагающегося в норме одного зародыша возникают два или больше зародышей, которые и называются однояйцевыми близнецами. В некоторых случаях раздвоение первоначально одного зародыша бывает неполное, или в одном яйце закладывается два зародыша, которые потом частично сливаются или срастаются, — тогда возникают так называемые «соединенные близнецы», «двойные уродства», «двойные образования» («Doppelbildungen»), особый подтип ОБ (см. главу шестую). В силу своего происхождения ОБ внутрипарно всегда одинакового пола: или оба мужского, или оба женского. ОБ вообще наследственно в высшей степени похожи, что обнаруживается в их сходстве по множеству признаков (рис. 1) и потому привлекает к ним интерес генетиков. Разнояйцевые близнецы (РБ), как показывает само название, в отличие от ОБ возникают из разных яиц, оплодотворенных разными спермиями. Их обычно считают братьями и сестрами, родившимися почти одновременно. В отношении пола РБ внутрипарно могут быть двух подтипов: или одинакового пола, или разного пола. Будем впредь условно первый подтип обозначать РБо (рис. 2), где буква «о» значит «одинакового пола», а второй подтип — РБр, где буква «р» значит «разного пола» (рис. 3). В смысле сходства пара РБ может быть столь же похожа или различна, как обычные братья-сестры (БС). Некоторые исследователи полагают, что только ОБ являются «истинными» близнецами, тогда как РБ считаются «ложными» близнецами (Caullery, 1945; Е. Wolff, 1948, и др.). Однако эта точка зрения имеет противников, думающих, что у обоих типов близнецов есть нечто специфически общее, как близнецов вообще, именно в биохимии обмена веществ, и что потому РБ нельзя считать вполне подобными братьям-сестрам (Gedda, 1951). Этот спор пока что остается неразрешенным. Изучение близнецов, помимо того, что дает ряд данных, интересных для теории биологии, также открывает пути для решения важных практических вопросов в области педагогики, медицины и животноводства, как это будет видно из соответствующих глав этой книги.


Глава первая
БЛИЗНЕЦЫ В МИФАХ, В КУЛЬТЕ, ФОЛЬКЛОРЕ, ХУДОЖЕСТВЕННОЙ ЛИТЕРАТУРЕ И ИЗОБРАЗИТЕЛЬНОМ ИСКУССТВЕ
Близнецы с древнейших времен вызывали к себе интерес вместе с изумлением и страхом, как явление необычное — ведь в человеческом обществе близнецы составляют около одного процента всех родов, а тройни и четверни или двойные уродства появляются еще более редко (см. главу шестую). О том, как воспринимались близнецы, что о них думали, как о них толковали древнейшие культурные народы, можно узнать из мифов, культов и обрядов, молитв, песен, сказок, легенд и т. д., а также из памятников изобразительных искусств (главным образом скульптуры). Так, в древнем Египте важнейшие божества — Осирис и Исида — были близнецы, ставшие супругами еще в материнской утробе. Главные божества древнего Ирана, бог света Ормузд и бог тьмы Ариман были близнецы-антагонисты. В Месопотамии у вавилонян и ассирийцев было несколько пар близнецовых божеств. Культ одной из этих пар, благодетельных божеств, был связан с созвездием, ныне называемым Близнецы. Вероятно, в астрономию это созвездие вошло впервые в связи с этим культом. У европейских народов созвездие Близнецов было соединено с культом древнегреческих близнецов — героев Диоскуров. В древнем Китае существовал культ близнецов Хэ-Хэ-эр-шань (Штернберг, 1927). В Индии, помимо других близнецовых божеств, существовал очень древний культ юных братьев-близнецов Асвинов, светоносных божеств утренней зари, благодетелей человечества (Миллер, 1876, и др.). Левины были целителями от разных недугов и болезней, покровителями домашних животных и т. д. В их имени звучит корень санскритского слова «лошадь» (asva), их представляли едущими на колеснице или верхом, укротителями и покровителями коней. Образ Асвинов очень близок эллинским Диоскурам, как мы увидим ниже.

В Элладе существовало множество местных культов близнецов, мифов и сказаний о них. Некоторые из этих близнецов вошли в общеевропейскую культуру в образах поэзии и искусства. Так, Аполлон (Феб) и Артемида (рис. 4), общеэллинские божества Солнца и Луны, были близнецами, детьми Зевса и богини Латоны. Другая пара — братья-близнецы Кастор и Поллукс (Полидевк) — герои спартанского происхождения. Но постепенно их культ стал достоянием всей древней Греции, а позже распространился и в Риме. Происхождение этих близнецов в разных сказаниях трактуется различно. Они — сыновья Леды, мужем которой был спартанский царь Тиндарей. Но Леду, как известно, пленил Зевс, явившись ей, по мифу, в образе лебедя; это изобразил на своей картине Леонардо да Винчи (рис. 5).
 От Зевса у Леды родилась знаменитая красавица Елена, героиня Троянской войны. По некоторым сказаниям, ее сестра Клитемнестра, жена Агамемнона и мать Ореста, была ее близнячка, но от смертного отца Тиндарея. Кастор и Поллукс, по Гомеру, оба — сыновья смертного отца, однако позже их обоих стали считать сыновьями Зевса, почему они получили прозвище Диоскуры (дети Зевса). Гораций называет их произошедшими «из того же яйца» (Сатиры, II. 1. 26—27). Такими их рисует Леонардо да Винчи на вышеупомянутой картине. Вторая пара близнецов на этой картине, вероятно, Елена Прекрасная и Клитемнестра. Наконец, возникло третье возможное объяснение происхождения Диоскуров: один из них, Кастор, был сыном смертного отца, а Поллукс — бессмертного, Зевса. Такое представление о происхождении близнецов от двух разных отцов встречается и у других народов. Быть может, в связи с этим миф приписывает близнецам известные различия, кроме бессмертия только одного из них. Кастор, например, был знаменит как укротитель коней, а Поллукс — как кулачный боец. Они — образец братской дружбы, жить в разлуке они не могли (Пиндар, Ода II). Их чтили как «благодетелей» людей, особенно как «спасителей» мореплавателей в бурю, как об этом говорится в одном из «Гомеровских гимнов» и у Феокрита в идиллии «Близнецы». Античные народы чтили Диоскуров, как уже говорилось, в образе созвездия Близнецов. Этих героев, похожих на Асвинов, представляли себе в виде «белоконных» всадников. Конниками изображали их на вазах и барельефах. Известны, например, гигантские античные статуи их в виде укротителей коней на площади Квирииала в Риме (рис. 6). Фигура одного из близнецов является почти полным зеркальным изображением другой. Уменьшенные мраморные копии этих статуй стоят у колоннады бывшего Конногвардейского манежа Кваренги близ Исаакиевского собора в Ленинграде (рис. 6). Они являются прообразами известных «Укротителей коней» Клодта на Апичковом мосту в Ленинграде. Так образы древних божеств стали основой для реалистических скульптур, украшающих Невский проспект.
От Зевса у Леды родилась знаменитая красавица Елена, героиня Троянской войны. По некоторым сказаниям, ее сестра Клитемнестра, жена Агамемнона и мать Ореста, была ее близнячка, но от смертного отца Тиндарея. Кастор и Поллукс, по Гомеру, оба — сыновья смертного отца, однако позже их обоих стали считать сыновьями Зевса, почему они получили прозвище Диоскуры (дети Зевса). Гораций называет их произошедшими «из того же яйца» (Сатиры, II. 1. 26—27). Такими их рисует Леонардо да Винчи на вышеупомянутой картине. Вторая пара близнецов на этой картине, вероятно, Елена Прекрасная и Клитемнестра. Наконец, возникло третье возможное объяснение происхождения Диоскуров: один из них, Кастор, был сыном смертного отца, а Поллукс — бессмертного, Зевса. Такое представление о происхождении близнецов от двух разных отцов встречается и у других народов. Быть может, в связи с этим миф приписывает близнецам известные различия, кроме бессмертия только одного из них. Кастор, например, был знаменит как укротитель коней, а Поллукс — как кулачный боец. Они — образец братской дружбы, жить в разлуке они не могли (Пиндар, Ода II). Их чтили как «благодетелей» людей, особенно как «спасителей» мореплавателей в бурю, как об этом говорится в одном из «Гомеровских гимнов» и у Феокрита в идиллии «Близнецы». Античные народы чтили Диоскуров, как уже говорилось, в образе созвездия Близнецов. Этих героев, похожих на Асвинов, представляли себе в виде «белоконных» всадников. Конниками изображали их на вазах и барельефах. Известны, например, гигантские античные статуи их в виде укротителей коней на площади Квирииала в Риме (рис. 6). Фигура одного из близнецов является почти полным зеркальным изображением другой. Уменьшенные мраморные копии этих статуй стоят у колоннады бывшего Конногвардейского манежа Кваренги близ Исаакиевского собора в Ленинграде (рис. 6). Они являются прообразами известных «Укротителей коней» Клодта на Апичковом мосту в Ленинграде. Так образы древних божеств стали основой для реалистических скульптур, украшающих Невский проспект.

Другого рода отзвук народных представлений об Асвинах-Диоскурах сохранился, по-видимому, в средневековой византийской и русской культуре. Под их влиянием сложились образы и «жития» близнецов-святых: Космы и Демьяна, Флора и Лавра (Миллер, 1876; Штернберг, 1916). Последние изображаются на иконах всадниками и почитаются как покровители коней и других домашних животных. Обе пары святых являются благодетелями рода человеческого, а Косма и Демьян в частности — врачами-бессребрениками, целителями от разных недугов, как Асвины. Множество других близнецовых пар фигурирует в античных мифах и сказаниях. Интересно, что некоторые из таких диад имеют резкие внутрипарные различия, т. е. различия между близнецами одной пары. Например, Геракл, сын Зевса и смертной Алкмены, родился с близнецом Ификлом, зачатым матерью от ее законного мужа Амфитриона. Незначительная фигура Ификла совершенно затмевается его могучим близнецом божественного происхождения. Но и дети-близнецы от Зевса и смертной могут быть резко различными людьми, как например Амфион и Зет, фиванские герои. Первый был кроткий, тихий человек и великий музыкант, тогда как его близнец Зет, в противоположность ему, был грубый и шумный силач, охотник. Их различие очень образно обнаружилось при постройке стен города Фивы: Зет с трудом и напряжением ворочал огромные камни, а Амфион,спокойно сидя, игрой на лире заставлял каменные глыбы двигаться и слагаться в стену. Эти близнецы изображаются с символическими атрибутами: Амфион с лирой, Зет с дубиной или охотничьей собакой. Оба героя являются также главными действующими лицами знаменитой античной скульптурной группы «Фарнезийский бык» (Неаполь), изображающей один из их подвигов. Другая пара близнецов-антиподов — боги-ветры: Борей (холодный) и Зефир (теплый) и т. д. Нет надобности останавливаться на остальных мифах об античных близнецах. Отметим лишь, что верховные божества Рима — Юпитер и Юнона — были близнецы. Основатель Рима, легендарный герой Ромул, как известно, имел близнеца-соперника Рема. Обоих вскормила волчица, как это многократно изображалось в скульптуре и живописи античных и новых времен (рис. 7). Из полулегендарных римских героев эпохи борьбы Рима с Альбой Лонгой Тит Ливий (I. XXIV) называет Горациев, трех братьев-близнецов, подвиг которых впоследствии воспел Корнель в своей трагедии «Гораций» (1640). Клятву их изобразил Давид на своем известном полотне (рис. 8).


У других европейских пародов в мифах и сказаниях также фигурируют близнецы. Например, в Эдде содержится история светлого бога солнца Бальдура и его слепого близнеца Гедура, убившего его. Близнецы здесь тоже антагонисты. Об этой паре сообщает еще Тацит («Германия», 43), сравнивая их с Диоскурами. В русской литературе гибель Бальдура, на основании скандинавских сказании, описал А. Майков в поэме «Бальдур». Быть может, и тройничные божества, как например Грации восходят в своем первоначальном образе к тройне. В культах и мифах, по-видимому, отразились также образы различных двойных уродств, поражавших воображение людей древности, например соединенные близнецы Молионы эллинской мифологии, имевшие две головы, четыре руки и четыре ноги, или двуликий римский бог Янус (рис. 9), именем которого называют теперь двойных уродов, имеющих два лица на одной общей голове (см. главу шестую). Сюда же относятся индийские демоны из свиты Мары, изображенные на барельефе одного храма, гавайский идол двойного женского божества (Martin, 1880) и другие.

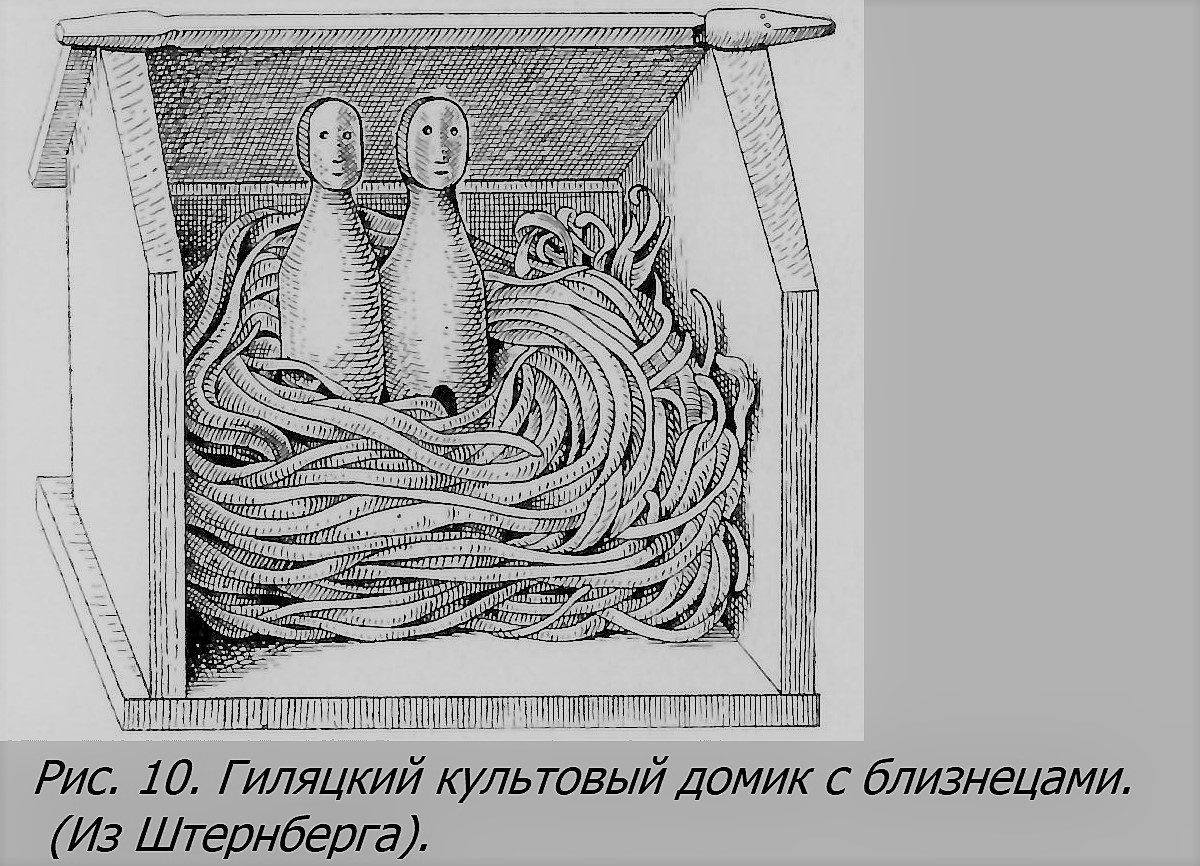
Обожествление близнецов происходило не только у древних народов. Оно наблюдается и у современных народов разных стран, стоящих на низкой ступени культуры, например в Сибири в XIX в. (гиляки, айны и др. Штернберг, 1916), в Африке (зулусы и др. — Брайант, 1953), в Южной Америке и т. д. Изучение причин обожествления близнецов у этих народов проливает некоторый свет на возникновение этого рода культа и в древности (Штернберг, 1916). По-видимому, здесь существенную роль играет то обстоятельство, как в донаучное время объяснялась причина появления на свет близнецов. Штернберг думает, что люди примитивной культуры, например гиляки, считали рождение близнецов явлением сверх-естественным. От каждого соития обыкновенных людей, полагали они, может родиться только по одному ребенку. А раз их родится одновременно два, то, кроме отца, в этом должен участвовать кто-то второй, т. е. женщина должна быть оплодотворена двумя разными лицами. Этим вторым может оказаться божество, доброе или злое. При предположении, что отцом второго близнеца является сверхъестественное лицо, в зависимости от того, благое это или злое божество, слагалось диаметрально противоположное отношение к близнецам и их матери. В первом случае близнецы были сами или боги, или герои, впоследствии обожествленные, как например Диоскуры, Геракл и другие. Интересно отметить, что влюбленный Зевс являлся к своим избранницам нередко в образе какого-нибудь животного, например к Леде, матери Диоскуров, в виде лебедя (рис. 5), а в других случаях в образе быка (к Европе) и т. д. Штернберг справедливо, по-видимому, усматривает в животных образах богов пережитки в греческих мифах древнего зооморфизма, обоготворения животных, столь распространенного особенно в древнем Египте. Так, волчица, кормящая Ромула и Рема, лишь в более поздних сказаниях оказалась их мамкой, по старинным же верованиям, она, вероятно, была их матерью, а отцом — волк; и лишь позже он был заменен богом Марсом, принимающим облик волка и похитившим их мать Рею Сильвию (silva — лес!). Человеческий образ божества лишь постепенно вытесняет звериный, и этот процесс отражается в двойственных представлениях многих сказаний и сказок. Зооморфическое представление о божестве недавно еще жило, например, среди гиляков Камчатки, которые считали, что «горный дух» является избранной им женщине в виде медведя и уносит её к себе в тайгу в качестве жены. Даже один администратор Камчатки начала прошлого века, человек из русской интеллигенции той эпохи, сообщает о таком факте, как достоверном, имевшем место якобы в Петропавловске (Штернберг, 1916). В связи с «божественным» происхождением близнецов гиляки имели до недавнего времени особый культ близнецов, с устройством специального святилища на месте их погребения, с особыми обрядами и т. д. К предметам близнецового культа у гиляков относятся и своеобразные домики с грубым изображение близнецов (рис. 10). Аналогичный культ близнецов описан у различных других современных народов, например в Африке у негров (Штернберг. 1916; Брайант, 1953, и др.) и т. д. Пережитки древней веры во вмешательство божества в дело рождения близнецов сохранилось еще и в некоторых цивилизованных странах. Например, в Перу еще недавно считалось, что бог молнии и дождя, названный в христианское время св. Яковом (Сантъяго), является повинным в рождении близнецов, почему их называли его именем, пока церковь во избежание соблазна не запретила этого. А в Англии, где существовало из древности идущее аналогичное представление, до последнего времени говорили относительно женщины, родившей близнецов: «Св. Мартин своим молотом ударил в ее окно». Этот святой заменил языческого бога-громовика (как у нас Илья — Перуна), молот которого олицетворял гром и молнию, последняя же была образом фаллоса (Штернберг, 1927).
При противоположном допущении, при вере в то, что вторым отцом одного из близнецов был злой дух, дьявол и т. д., близнецов убивали или убивали одного из них, считавшегося именно ребенком злого духа, иногда преследовали и убивали и мать, совершали очистительные обряды и т. д. Интересно, что такое резко различное отношение к близнецам встречалось у племен примерно одного культурного уровня и живущих даже сравнительно недалеко друг от друга, например у разных племен зулусов и других народностей (Штернберг, 1916; Hall, 1928; Брайант, 1953, и др.). Вопрос о происхождении близнецов тесно связан с другим, очень характерным для их понимания — вопросом о сходстве и различии близнецов. По представлениям различных древних народов, близнецы могли резко отличаться друг от друга, даже будучи одного пола: те же Диоскуры и другие пары близнецов древних греков. Внешнее различие, разница способностей и т. д. только в некоторых случаях зависят от различия отцов. Это различие иногда сочетается с большой взаимной любовью, как у Диоскуров; иногда же — с соперничеством и взаимной враждой, как у Исава и Иакова, по библии, Ромула и Рема и т. д. В этих случаях, очевидно, в мифах и легендах были РБ, так как близнецы заметно различны. Но в ряде случаев речь идет о чрезвычайном сходстве близнецов, очевидно ОБ, из-за чего происходит ряд ошибок — их принимают одного за другого. Это сходство близнецов — неразрешимая гадка для ума примитивного человека и объясняется только волшебством. Так и говорится в одной древнегреческой сказке о братьях-близнецах (Зелинский, 1922), легшей в основу целой серии комедий разных эпох; из них самой древней из сохранившихся является известная римская комедия Плавта «Менехмы». На основе ее написал свою «Комедию ошибок» Шекспир, а далее возник ряд комических пьес, в том числе комедии Гольдони, зингшпиль «Близнецы» Шуберта и другие. Сходство близнецов является причиной различных забавных ошибок и путаницы. Мотив дружбы обычно чрезвычайно похожих братьев-близнецов, дружбы, заставляющей одного из них разыскивать и выручать другого, лежащий в основе «Менехмов», встречается с различными вариантами в сказках и других европейских народов, например в русской сказке «Два Ивана солдатских сына» (сб. Афанасьева), немецкой — «Два брата» (Братья Гримм), эстонской «О быстроногом, леворуком и зоркоглазом» (Крейцвальд), где фигурирует тройня, и т. д. Близнецы как литературный сюжет встречаются не только в комедиях, но и в различных других жанрах — повестях, романах и т. д., то как основная, то как побочная тема (см.: Poll, 1930; Gedda, 1951, и др.). Взаимоотношения близнецов и ситуации, в которые они попадают, оказываются весьма разнообразными и далеко не всегда комичными. Так, сходство близнецов не всегда является гарантией их дружбы. Наоборот, на почве его может возникнуть ненависть, более глубокая, возможно, чем при наличии различий. Такую ненависть рисует Андерсен-Нексе в новелле «Последыши». В более абстрактной форме о такой ненависти говорит Тургенев в стихотворении в прозе «Близнецы». Художественная литература затрагивает в таких случаях вопросы, еще не разработанные наукой, и порой очень верно, черпая из жизни, изображает их. Интересна, например, попытка Жорж Санд в повести «Маленькая Фадетта» изобразить обусловленные отчасти обстоятельствами жизни различия психики близнецов, вероятно ОБ, оказавшихся в сложной ситуации, когда оба были влюблены в одну и ту же девушку, предпочитающую одного из них. Пожалуй, еще более интересным примером художественного описания зависимости психики близнецов от условии среды может служить малоизвестная повесть Тараса Шевченко «Близнецы». В ней описана жизнь двух чрезвычайно похожих в детстве мальчиков-близнецов, явно ОБ, которые дальше пошли совершенно разными жизненными путями. Один стал офицером, пьяницей и негодяем, другой — врачом, благородным и гуманным человеком. Шевченко очень правдиво изображает «воспитывающую» роль социальной среды и показывает, как из неразличимо-похожего исходного материала — детей-близнецов — в разных условиях жизни образуются совершенно различные люди. Наука еще не исследовала достаточно глубоко того, что в данном случае показала талантливая повесть. Уже в мифах и сказках ясно выступает представление о том, что близнецы могут быть разного пола (Аполлон и Артемида, Осирис и Исида и т. д.), что они могут оказаться, будучи одинакового пола, очень различными (Амфион и Зет, Исав и Иаков, Ормузд и Ариман и т. д.). Здесь речь идет несомненно о РБ. В тех же случаях, где сходство близнецов столь велико, что их не различает даже жена одного из них, как в сказке «Два Ивана» и других вышеупомянутых произведениях, тут явно имеются в виду ОБ. Подобный материал черпался несомненно из жизни, из множества занятных историй и анекдотов. В прошлом веке о таких рассказах писал Гальтон (Galton, 1875), и до наших дней они служат для развлечения, как например в произведениях Марка Твэна или в стихах Маршака (1954). Причины сходства и различия близнецов, их природа и происхождение, как видно из вышеизложенного, на заре истории уже начавшие интересовать людей, с появлением и развитием науки стали предметом ее изучения. К краткому обзору развития науки о близнецах мы теперь и обратимся.
Глава вторая
ИЗ ИСТОРИИ НАУКИ О БЛИЗНЕЦАХ
Научное изучение близнецов началось в древней Греции. Сохранились указания на то, что еще в V в. до н. э. философы Эмпедокл и Демокрит высказывались относительно происхождения близнецов. Оба считали, что близнецы получаются от избыточного семени. В разных вариантах эта мысль повторялась в римскую эпоху (Гален, II в. н. э. и др.) и в средние века. Гиппократ, «отец медицины», в приписываемой ему книге «О семени и природе ребенка» говорит, что близнецы рождаются от одного «семени» (понимаемого тогда как жидкость), которое попадает в разные «пазухи» матки; этим определенные дозы семени оказываются изолированными друг от друга, что и ведет к образованию разных зародышей поблизости друг от друга — близнецов. Другая гипотеза заключалась в том, что близнецы образуются благодаря оплодотворению разными отцами (Эразистрат, около III в. до н. э. и др.) — мысль, как мы уже видели, известная еще из мифов и повторявшаяся до наших дней. Наибольшее научное значение имели воззрения Аристотеля (IV в. до н. э.). О близнецах он говорит несколько раз в своем сочинении «О возникновении животных». Он указывает на связь уродств с многоплодием, особенно у птиц, в частности у кур, в яйце которых «зачатки, лежащие близко друг от друга, срастаются». «Уродства, - пишет Аристотель, — чаще встречаются у многородящих. Поэтому у человека их меньше... но в местностях, где женщины многоплодны, уродства случаются чаще, например в Египте». Он пытается установить причину возникновения близнецов, усматривая аналогию между возникновением избыточных органов и близнецами: «... а что касается причины возникновения излишних частей, то она такова же, как и рождение близнецов. Причина эта лежит уже в зачатках; если материи сгустится больше, чем следует по природе части, тогда именно случится, что одна часть будет больше других, например палец, рука или какой-нибудь другой из конечных членов, или же вследствие расщепления зачатка их возникает несколько, подобно водоворотам в реках. Ибо и в них, если текущая и обладающая особым движением вода наталкивается на какое-нибудь препятствие, то из одного вихря образуются два с тем же самым движением; то же происходит и с зачатками» (Кн. 4, гл. 4). Мысль о расщеплении зачатка и динамическое представление о нем в сравнении с водоворотом удивляет своим сходством с представлениями XX в., основанными на значительном фактическом материале (Graper, 1931; Newman, 1923—1948, и др.). Замечательно рассуждение Аристотеля о сущности уродства: «Уродство принадлежит к числу необыкновенных явлений, совершающихся, однако, не вопреки всей природе, а вопреки лишь большинству природных явлений» (Кн. 4, гл. 3). Борясь с суеверными представлениями своей эпохи об уродствах, а также о близнецах, Аристотель стремится весь вопрос поставить на научную почву, что удалось сделать лишь через два тысячелетия после него. В средние века, когда в течение столетий биологические сочинения Аристотеля были утрачены и забыты, о близнецах и двойных уродствах царили самые нелепые взгляды, матери таких детей обвинялись в сношениях с нечистой силой или зверями, жестоко преследовались и т. д. Много фантастических представлений по этим вопросам сохранилось и в сочинениях эпохи Возрождения (Colombo, 1559; А. Раге, 1578, и др.). Даже в работах XVII в., например у Лицетуса (Licetus, 1616, 1665), наряду с интересными фактами, взятыми из действительности, как известный генуезец Коллоредо с близнецом-паразитом (рис. 11), встречаются описания фантастических двойных уродов, заимствованные из разных рассказов (рис. 12).


Но уже в середине XVII в. появилась книга Гарвея «О зарождении животных» (Harvey, 1651), в которой автор, возвращаясь к аристотелевской точке зрения на развитие зародыша (эпигенетическое), на основании собственных наблюдений в области эмбриологии развивает новые научные взгляды, являясь основателем эмбриологии нового времени. И хотя о близнецах он в общем высказывается мало, в духе Аристотеля, все же для дальнейшего изучения этого вопроса труд Гарвея сыграл большую роль. В XVIII в. интерес к различным уродам — «монстрам», — в частности двойным, был Очень велик. Петр I разделял его, и по его указам у нас в России собирались различные уродства, в том числе живые уроды, которые помещались в первом русском музее в Петербурге — Кунсткамере (Бэр, 1850; Станюкович, 1953). Коллекции двойных уродств этого музея впоследствии изучали Вольф, Бэр, Загорский, Яцута и другие. С развитием теории преформации вопрос о близнецах, особенно соединенных, и вообще об уродствах стремятся решить так же на основе этой теории. Слагаются два взгляда, борьба которых развертывается в течение 20 лет (1724—1743 гг.)в Парижской Академии наук — это знаменитый «спор о монстрах» (Marlin, 1880). Можно предположить, согласно теории преформации, что первоначально заложенное в яйце строение зародыша нарушается по разным причинам — тогда возникает уродство; двойной же урод возникает путем слияния двух зародышей. Эту точку зрения в упомянутом споре развивал Лемери (Lemery). Против него выступал Винсло (Winslow), считавший, что в яйце уже изначально имеется раздвоенный зародыш или уродство. Спор потерял свою остроту и смысл с развитием новых точек зрения на основе новых фактов. Большую роль в этом отношении сыграли труды К. Ф. Вольфа, развивавшего эпигенетический взгляд на эмбриогенез в своей «Теории развития» (1759). Вольф, отвергая учение о слиянии двух зародышей при образовании близнецов, считал, что близнецы возникают благодаря «избыточному развитию», идущему в яйце в разных направлениях. Он изучал (1773) близнецов в курином яйце, показав их развитие из одной яйцеклетки (рис. 13), а также соединенных близнецов («двухголового теленка»), доказывая, что последние возникли не путем срастания двух зародышей, а раздвоением одного.

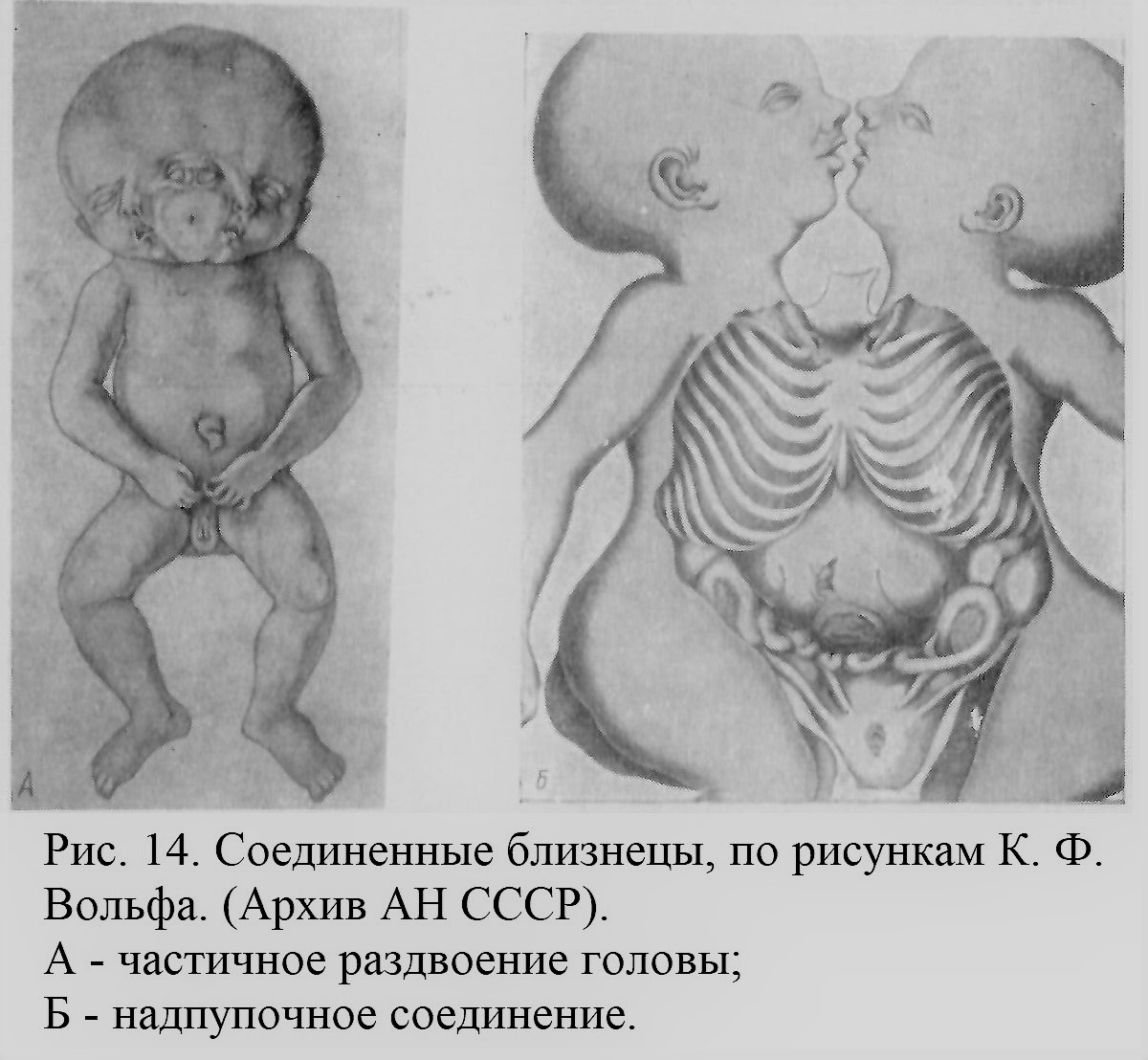
Позже, живя в Петербурге, он много потрудился над изучением анатомии соединенных близнецов Кунсткамеры (рис. 14). Труд этот остался незаконченным из-за смерти Вольфа. Лишь небольшая часть этого обширного материала, хранящегося в Архиве Академии наук СССР, была в свое время опубликована (1780; ср. Райков, 1952; Бляхер, 1955).
Взгляды Вольфа развивал дальше Меккель (Meckel, 1815). Он считал, что правая и левая половины зародыша возникают самостоятельно и могут оставаться в той или иной мере несоединенными; в таком случае недостающие части образуются у них заново, как у гидры при регенерации ее. Причиной такого раздвоения зародыша Меккель считал изменение «направления и энергии образующей силы» под влиянием различных обстоятельств. Им высказана плодотворная мысль о задержке развития как о стимуле к возникновению близнецов. Взгляды Вольфа и Меккеля об эпигенетическом происхождении близнецов из одного яйца развивал дальше крупнейший эмбриолог первой половины XIX в. К. М. Бэр. Создатель сравнительной эмбриологии на основе учения об единстве типа позвоночных, Бэр пользовался сравнительным методом и при изучении близнецов. Он описал «двойной эмбрион курицы» (1827) на ранней стадии развития и позже сравнивал его с двумя экземплярами двойного уродства переднего раздвоения (рис. 15) у рыб (Бэр, 1845а). Бэр, подобно Вольфу, считал, что они являются результатом расщепления первоначально единого эмбриона, а не слияния двух эмбрионов, как подобные явления склонны были толковать его современники, оба Жоффруа Сент-Илера. Прежде чем перейти к последним, надо отметить, что Бэр много лет занимался соединенными близнецами (Канаев, 1951), детально изучал их анатомию, в частности интересовался соединенными головами человеческими близнецами (краниопаги, — рис. 50) и даже пытался исследовать физиологию пары таких живых младенцев (1856).

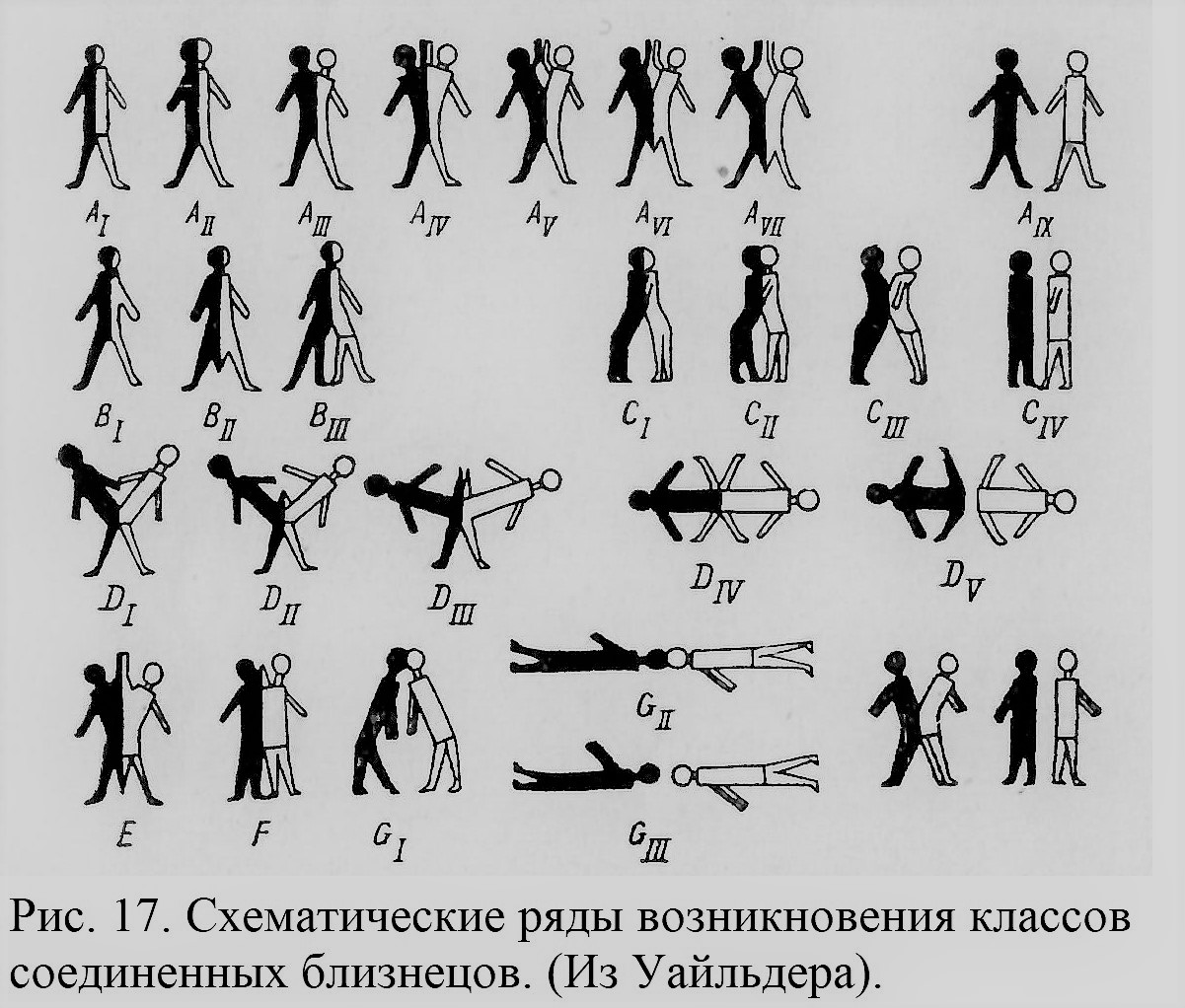
Идея единства типа, восходящая к Аристотелю, в конце XVIII в. в работах Гёте, а позже У. Жоффруа Сент-Илера поднялась на новую ступень развития благодаря разработке сравнительной анатомии с помощью принципа гомологий. Для Гёте роды, виды, особи являются метаморфозами одного общего «типа», т. е. видоизменениями под влиянием условий среды. Единство типа и постепенность метаморфозов в чисто морфологическом смысле хорошо демонстрировались путем построения рядов метаморфозов. Под влиянием учения Гёте такой ряд для двойных уродств (рис. 16) составил анатом Зёммеринг (Sommering, 1791). Впоследствии подобные ряды, показывающие постепенный переход раздвоения определенного рода от одной степени к другой, успешно разрабатывались, например, Уайльдером (Wilder, 1904; см. рис. 17) и другими авторами (Schwalbe, 1907, 1923; Potter, 1953).

Этьенн Жоффруа Сент-Илер (Е. Geoffroy Saint-Hilaire, 1822), известный поборник идеи единства типа, стремился, между прочим, экспериментально показать возможность появления видоизменений формы под влиянием воздействия среды. Сам Этьенн Жоффруа Сент-Илер решающих результатов в этой области не получил, но его идеи развивал дальше его сын Исидор (1837) и более поздние последователи, особенно Дарест (Dareste, 1891). Исидор Жоффруа Сент-Илер с небывалой до того полнотой и последовательностью изучил и систематизировал огромный материал по разнообразным уродствам, являясь основателем тератологии, науки об уродствах. Третья часть его обширного труда посвящена соединенным близнецам. Система Исидора построена на идее единства типа. Его классификация и терминология частично живет в современных работах, особенно французских ученых (ср. Е. Wolff, 1948, и др.). Эта система стала отправным пунктом всех дальнейших работ систематизаторов двойных уродств — Маршана (Marchand,1897), Уайльдера (Wilder, 1904),Швальбе (Schwalbe, 1907), Поттер (Potter, 1953). Сент-Илеры считали соединенных близнецов, вопреки Вольфу и Бэру, результаты срастания двух зародышей, по, по-видимому, возникающих в одном яйце и имеющих общие оболочки. Следуя своему отцу, Исидор Сент-Илер считал, что слияние близнецов происходит по принципу «сродства подобного с подобным», согласно которому соединяются тождественные части: сердце с сердцем, позвоночник с позвоночником и т. д. Эта идея, без объяснения причин такого явления, недавно нашла поддержку (см. главу шестую (окончание)). Дарест был одним из основоположников экспериментальной тератологии, восходящей еще к работам Реомюра (1751), особенно же к работам Сент-Илеров. Дарест выяснил, при каких условиях получаются соединенные близнецы, т. е. образование двух эмбрионов в одном яйце на достаточно близком расстоянии друг от друга. Однако экспериментально близнецов ему получить не удалось. Это сделали в самом конце XIX в. Дриш (Driesch, 1891), Гертвиг (Hertwig, 1893), Шульце (Schulze, 1894), Леб (Loeb, 1894) и многие другие, особенно же успешно в начале XX в. этот вопрос разработали на амфибиях Шпеман и его школа . Так, на грани XIX и XX в. наступила эпоха экспериментального изучения близнецов, сменившая чисто анатомическое и эмбриологическое описание их в XVIII в. и первой половины XIX в. В качестве опередивших в свое время пионеров экспериментального изучения образования близнецов следует упомянуть Якоби (Jacobi, 1765), впервые описавшего возникновение близнецов и двойных уродств у рыб при искусственном оплодотворении икры, Валентина (Valentin, 1851), пытавшегося продольным разрезом получить из куриного эмбриона близнецов, что ему удалось на одном экземпляре (заднее раздвоение), Кноха (Knoch, 1873), получившего близнецов у рыб путем сотрясения воды с оплодотворенной икрой и использования других «вредных» условий; правда, еще Валентин знал, что от тряски при перевозке икры в воде увеличивается число близнецов. В связи с успехами в области экспериментального изучения эмбриогенеза близнецов, микроскопического изучения половых клеток и оплодотворения к концу XIX в. уже достаточно отчетливо выяснилось представление о двух типах близнецов ОБ и РБ, которое лишь в гипотетической форме было высказано в 1874 г. Дарестом и приблизительно в то же время другими исследователями. Установление этих двух типов близнецов шло также еще другим путем: со стороны акушеров, изучавших человеческих близнецов (см.: Kleinwatchter, 1871, и др.). Исследование оболочек, хориона и амниона, а также плаценты указывало, что есть близнецы с одним общим хорионом и есть с разными хорионами; первых стали считать за ОБ, вторых за РБ. Только недавно была внесена существенная поправка в это представление о связи оболочек с типом близнецов (см. главу четвертую). Различение двух типов близнецов дало возможность поставить вопрос о большом методическом значении изучения близнецов для генетики. Это сделал двоюродный брат Ч. Дарвина антрополог Гальтон (Galton, 1875; более правильная транскрипция — Голтон). Считая пару ОБ наследственно одинаковой, Гальтон думал, что внутрипарные различия между близнецами надо приписать влиянию факторов среды. Таким образом, близнецы могут служить средством для изучения вопроса о взаимоотношении наследственности и среды, «природы и питания» («nature and nurture»), как выражался Гальтон. Так возникла идея «близнецового метода», являющегося в наше время ценным методом для изучения генетики различных организмов, в частности человека (см. главу пятнадцатую). Но Гальтон не умел достаточно отчетливо различать оба типа близнецов, что было помехой для успешного развития его метода как в руках его самого, так и его ближайших последователей (Thorndike, 1905; Poll, 1914, и др.). Только в 1924 г. Сименсом (Siemens) был предложен новый метод, «метод сходства», для диагноза типа близнецов, дающий возможность выяснить, является ли данная пара близнецов одно- или двух-яйцевой. Этот метод был развит и усовершенствован другими учеными (Фершюр, Ньюмен и др.). Диагноз ставится путем сравнения у близнецов определенной группы признаков — цвета волос, кожи, глаз, группы и факторов крови, отпечатков кожных узоров пальцев и ладоней, формы лица, уха и т. д. ОБ по этим признакам внутрипарно более похожи, чем РБ. Существующие варианты этого метода диагноза близнецов дают возможность почти безошибочно решать вопрос, является ли данная пара ОБ или РБ. Это раскрыло широкие возможности для развития и использования близнецового метода. Изучение близнецов, особенно в генетическом аспекте, наиболее успешно велось последнее время в Германии (Сименс, Фершюр и его школа, Люксенбургер, Кронахер и др.), в США (Ньюмен, Райф, Гизелл, Коллман и др.), далее в Швеции (Дальберг), Голландии (Варденбург), Италии (Гедда), Японии (Комаи) и других странах. В СССР интенсивная работа была развита Медико-генетическим институтом в Москве; после его закрытия работа с близнецами у нас почти вовсе прекратилась. Интерес к близнецам и научным вопросам, с ними связанным, имеет известный общественный отклик как в европейских государствах, так и в других странах. О близнецах пишут статьи и книги для широких кругов читателей, создаются кинофильмы, организуются клубы и съезды близнецов (рис. 18).

Близнецы у растений стали изучаться только в связи с развитием микроскопии, преимущественно с XIX в. у них установлены те же два основных типа, ОБ и РБ, как и у животных; однако в силу своеобразия растительных организмов идентификация основных типов близнецов у них с этими типами у животных встречает ряд трудностей и вызывает споры. Переходя к современному состоянию науки о близнецах, мы прежде всего остановимся на некоторых общих вопросах, далее перейдем к близнецам у животных и затем у человека. Поскольку вопрос о близнецах у растений остается в стороне от основного развития темы этой книги, он вынесен в приложение, в конец книги .
Глава третья
ФАКТОРЫ МНОГОПЛОДИЯ
Прежде чем обратиться к рассмотрению вопроса о непосредственных причинах возникновения близнецов, надо познакомиться с некоторыми условиями, влияющими на процесс многоплодия. Такие условия, которые можно назвать факторами многоплодия, далеко еще не достаточно полно и разносторонне изучены. Мы кратко остановимся только на некоторых из них, относительно которых в литературе есть уже в той или иной мере разработанный материал, дающий основания для определенных обобщении и выводов. Возраст матери несомненно сказывается на рождении близнецов: частота таких родов до известного возраста матери увеличивается. Впервые связь числа предшествующих беременностей и возраста матери с количеством близнецовых родов установил, по-видимому, Дункан (Duncan) в 1865 г., почему некоторые авторы эту зависимость называют «законом Дункана». Что касается первого условия «закона Дункана» — числа предшествующих беременностей, то этот вопрос еще мало исследован. С возрастом в общем увеличивается и число предшествующих беременностей. Недавно была сделана попытка разделить эти два «фактора» путем сравнения первородящих матерей разных возрастов с матерями тоже разных возрастов, но рожавших многократно, вычислив проценты близнецовых родов у того и другого ряда. Оказалось, что между теми и другими матерями в отношении близнецовых родов разницы нет, иначе говоря, многорожавшая мать имеет такие же шансы родить близнецов, как и первородящая (Zazzo, 1955). Этот вопрос надо отличать от вопроса о матерях, повторно рожающих близнецов (см. далее в главе). В отношении же влияния возраста матери на рождение близнецов «закон Дункана» неоднократно проверялся и был подтвержден на большом статистическом материале, хотя из-за различия материала и, вероятно, по другим обстоятельствам разные авторы определили несколько различно возраст матери, на который падает максимум родов двоен: 26— 30 (Rumpe, 1891); 28.6 (Hirt, 1902), и т. д. В качестве примера рассмотрим цифры (в %) рождения близнецов во Франции за 1907—1910 гг. (Dahlberg, 1926) (табл. 1).
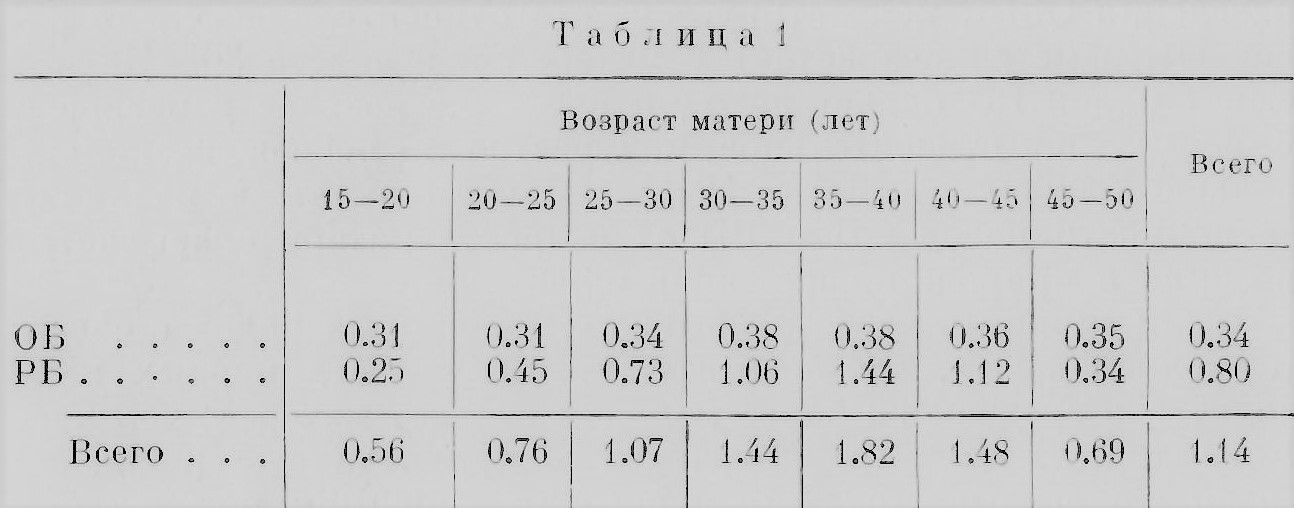

Эти цифры становятся наглядными, будучи выражены кривыми (рис. 19). Бросается в глаза различие в изменении с возрастом матери относительного количества родов РБ и ОБ. Число родов РБ заметно растет до 37—38 лет, а затем начинает круто падать. Число же родов ОБ почти не меняется, и после 35 лет можно отметить лишь очень незначительный подъем. Эти данные недавно подтвердил Стокс (Stocks, 1952) на статистических материалах Англин и Уэльса за 1943—1947 гг. Однако в одном исследовании, проведенном на негритянском населении США, констатируется, что относительное число родов ОБ заметно возрастает у матерен в возрасте 30—40 лет. «Пик» родов РБ падает на один и тот же возраст у негров и убелых, но относительное количество у негров несколько больше (Enders a. Stern, 1948). Возможно, что в различии зависимости родов ОБ и РБ от возраста матери сказывается разница в природе обоих типов близнецов. Если сравнить частоту обычных, одиночных родов с частотой близнецовых, то оказывается, что частота первых приходится в среднем на более молодой возраст матерей, чем частота вторых. Если же к сравнению привлечь еще рождение троен, то оказывается, что в среднем максимум родов троен приходится на еще более поздний возраст матерен, чем рождение двоен. Гедда (Gedda, 1951) приводит цифры для итальянского населения за 1930—1937 гг., выражая относительные числа родов в процентах; мы заимствуем лишь итоговые средние числа (табл. 2).
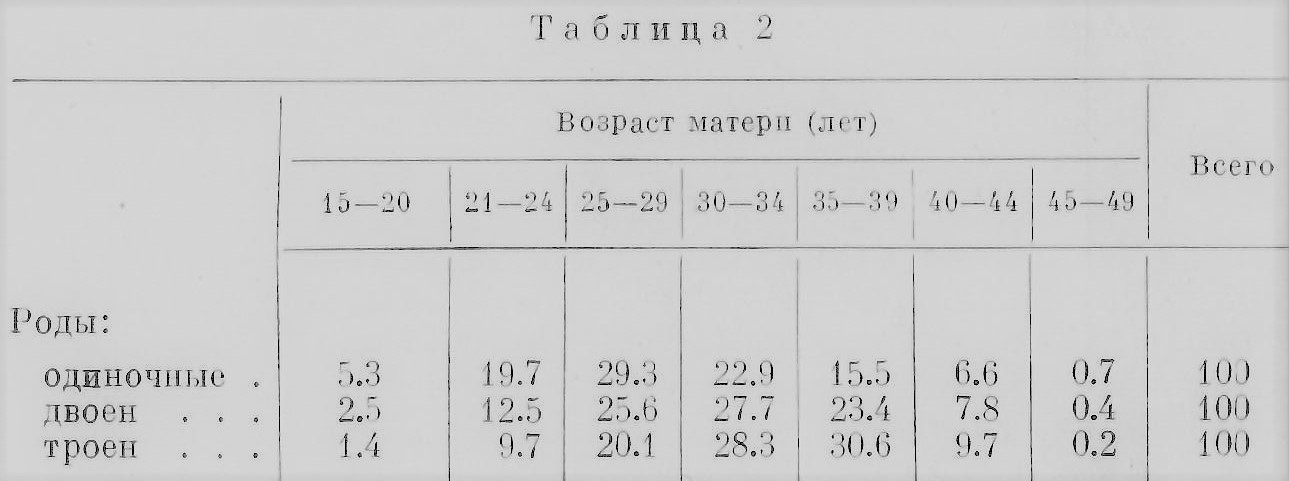 Мы видим, что максимум родов одиночек падает на возраст 25—29 лет, максимум двоен — на 30—34 года, а троен на 35— 39 лет. Гедда это наблюдение обобщает, говоря, что чем больше число новорожденных в одни роды, тем больше максимальная частота таких родов смещается на более поздний возраст матери. Это обобщение требует, конечно, проверки. На финском материале, недавно опубликованном (Miettinen, 1954), «принцип» Гедды на тройнях подтверждается: большинство матерей троен имело возраст 35—39 лет, в среднем 33 года, в общем они были заметно старше средней массы матерей. По для матерей четверен (правда, таких матерей в данном материале было только 18) повышения возраста установить не удалось: в среднем он был 32.5 года, т. е. даже несколько меньше, чем возраст матерей троен.
По данным Дальберга (Dahlberg, 1951), «пик» рождения троен приходится примерно на тот же возраст, как и двоен, — между 35—40 годами, но процент троен у более старых матерей примерно в 10 раз больше, чем у молодых, тогда как для двоен такой процент возрастает всего в 3—4 раза.
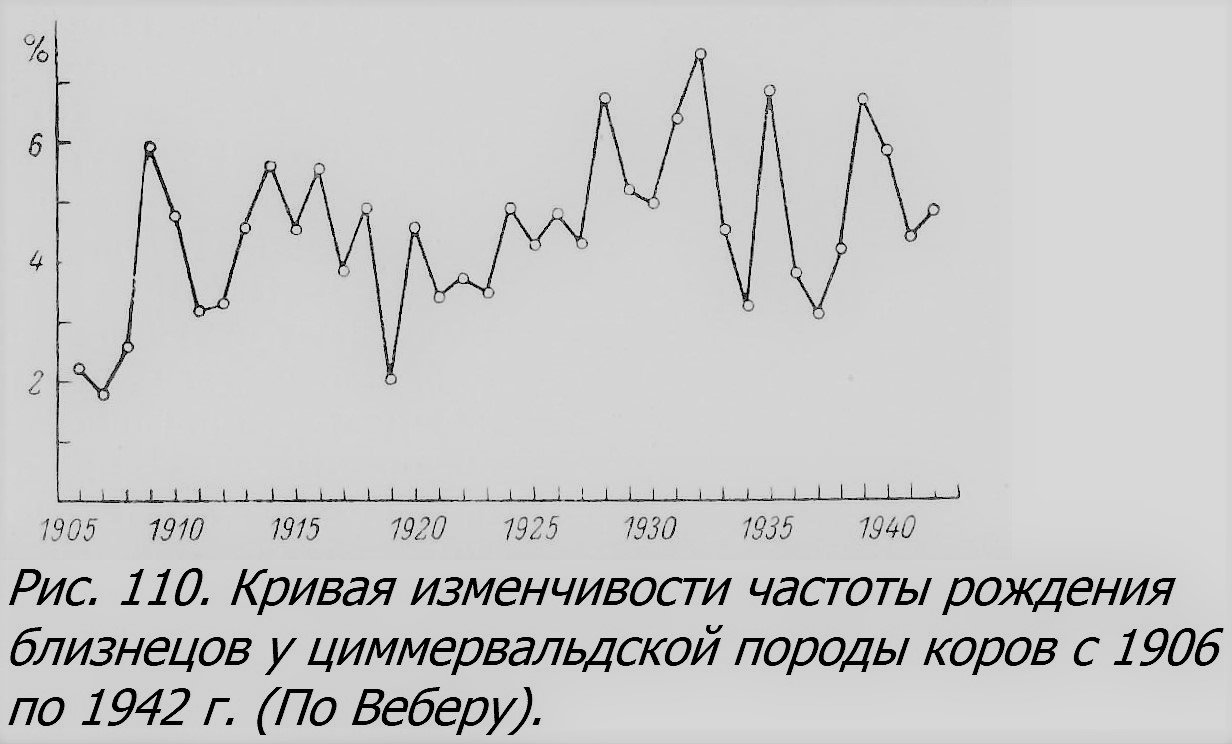
У животных влияние возраста матери на многоплодие сравнительно еще мало изучено и лучше всего известно у домашних животных — овец, свиней, коров и других (McArthur, 1942). Так, например, у коров процент близнецов возрастает с числом отелов, т. е. с возрастом животного, до 8—9-й беременности, после чего происходит перелом и процент уменьшается (рис. 110, Weber, 1945).
Мы видим, что максимум родов одиночек падает на возраст 25—29 лет, максимум двоен — на 30—34 года, а троен на 35— 39 лет. Гедда это наблюдение обобщает, говоря, что чем больше число новорожденных в одни роды, тем больше максимальная частота таких родов смещается на более поздний возраст матери. Это обобщение требует, конечно, проверки. На финском материале, недавно опубликованном (Miettinen, 1954), «принцип» Гедды на тройнях подтверждается: большинство матерей троен имело возраст 35—39 лет, в среднем 33 года, в общем они были заметно старше средней массы матерей. По для матерей четверен (правда, таких матерей в данном материале было только 18) повышения возраста установить не удалось: в среднем он был 32.5 года, т. е. даже несколько меньше, чем возраст матерей троен.
По данным Дальберга (Dahlberg, 1951), «пик» рождения троен приходится примерно на тот же возраст, как и двоен, — между 35—40 годами, но процент троен у более старых матерей примерно в 10 раз больше, чем у молодых, тогда как для двоен такой процент возрастает всего в 3—4 раза.
У животных влияние возраста матери на многоплодие сравнительно еще мало изучено и лучше всего известно у домашних животных — овец, свиней, коров и других (McArthur, 1942). Так, например, у коров процент близнецов возрастает с числом отелов, т. е. с возрастом животного, до 8—9-й беременности, после чего происходит перелом и процент уменьшается (рис. 110, Weber, 1945).
Существует еще одна особенность родов близнецов, зависящая от матери, на чем надо остановиться. Оказывается, что мать, однажды родившая близнецов, обнаруживает «тенденцию» к повторным родам таковых (Dahlberg, 1952). Хотя повторные роды идут параллельно с увеличением возраста матери, все же отмеченную Дальбергом «тенденцию», по-видимому, нельзя полностью сводить на возрастное явление и не замечать в ней определенную особенность, объяснение которой Дальберг видит в известном расстройстве специального гормонального механизма, о котором речь будет дальше. Дальберг, на основании данных Генеалогического бюро в Упсале, приводит следующий материал по этому вопросу (табл. 3), подтверждающий его прежние выводы (Dahlberg, 1926).
 Из табл. 3 явствует, что процент повторных родов близнецов (3.63) выше средней рождаемости близнецов в данной группе населения (популяции). Вычисленные по методу Вейнберга (см. главу девятую) количества РБ и ОБ показывают, что первых оказывается 4.55%, а вторых 1.43%. Повышение рождаемости касается обоих типов близнецов, причем цифра для РБ статистически достоверна, а для ОБ не достоверна, так как утроенная средняя ошибка ( +/- 0.90) превышает эту цифру (1.43). Стокс (Stocks, 1952) на основании новых данных считает, что для матери, имевшей близнецов, шансы иметь таковых повторно в 10 раз больше, чем для средней массы женщин.
Дальберг пытается дать физиологическое объяснение этого явления. Он думает, что в основе его лежит известное нарушение обычного гормонального механизма, регулирующего овуляцию. Дальберг основывается на опытах Цондека и Ашгейма, а также других, показавших, что гормон гипофиза стимулирует созревание яиц. С другой стороны, созревание лишь одного яйца у женщины регулируется тем, что, когда среди созревающих яиц одно из них достигнет известной степени зрелости, из его фолликула выделяется гормон фолликулин, задерживающий созревание других яиц. Если выделение гормона запаздывает или замедляется, то может успеть созреть и второе яйцо, что и поведет к образованию РБ.
Какие гормоны участвуют в этом сложном явлении (фолликулин оказывается смешанного состава), как оно связано с возрастом и т. д. — все это вопросы, пока еще недостаточно выясненные.
Рассмотренное объяснение применимо, однако, только в отношении РБ и то лишь для части случаев. Для возникновения ОБ надо предположить другой механизм, о котором речь будет ниже.
М. М. Завадовский (1941), исходя из тех же опытов с гонадотропными гормонами гипофиза, как и Дальберг, разработал метод для стимуляции многоплодия у овец, пригодный и для других домашних животных. Этот метод нашел применение в сельском хозяйстве. Для массовой работы с овцами бралась сыворотка крови жеребой кобылы (СЖК), содержащая гормон гипофиза; инъекция этой сыворотки за 2—5 дней до естественной овуляции увеличивала продукцию яиц почти вдвое, что выражалось в значительном учащении случаев многоплодия, как это, например, показывает диаграмма, обобщающая работу над отарой овец в одном из совхозов Средней Азии (рис. 20). Приплод благодаря методу Завадовского в среднем увеличивается на 50 и более ягнят на каждые 100 маток. В некоторых случаях вместо 1—2 ягнят в контрольной группе подопытные овцы приносили по 5 и 6 за раз. Методом Завадовского многоплодие, очевидно, достигается благодаря образованию РБ. Физиологический механизм, на котором основан метод, в основном ясен, хотя для ряда деталей вопроса требуются еще дополнительные исследования.
Из табл. 3 явствует, что процент повторных родов близнецов (3.63) выше средней рождаемости близнецов в данной группе населения (популяции). Вычисленные по методу Вейнберга (см. главу девятую) количества РБ и ОБ показывают, что первых оказывается 4.55%, а вторых 1.43%. Повышение рождаемости касается обоих типов близнецов, причем цифра для РБ статистически достоверна, а для ОБ не достоверна, так как утроенная средняя ошибка ( +/- 0.90) превышает эту цифру (1.43). Стокс (Stocks, 1952) на основании новых данных считает, что для матери, имевшей близнецов, шансы иметь таковых повторно в 10 раз больше, чем для средней массы женщин.
Дальберг пытается дать физиологическое объяснение этого явления. Он думает, что в основе его лежит известное нарушение обычного гормонального механизма, регулирующего овуляцию. Дальберг основывается на опытах Цондека и Ашгейма, а также других, показавших, что гормон гипофиза стимулирует созревание яиц. С другой стороны, созревание лишь одного яйца у женщины регулируется тем, что, когда среди созревающих яиц одно из них достигнет известной степени зрелости, из его фолликула выделяется гормон фолликулин, задерживающий созревание других яиц. Если выделение гормона запаздывает или замедляется, то может успеть созреть и второе яйцо, что и поведет к образованию РБ.
Какие гормоны участвуют в этом сложном явлении (фолликулин оказывается смешанного состава), как оно связано с возрастом и т. д. — все это вопросы, пока еще недостаточно выясненные.
Рассмотренное объяснение применимо, однако, только в отношении РБ и то лишь для части случаев. Для возникновения ОБ надо предположить другой механизм, о котором речь будет ниже.
М. М. Завадовский (1941), исходя из тех же опытов с гонадотропными гормонами гипофиза, как и Дальберг, разработал метод для стимуляции многоплодия у овец, пригодный и для других домашних животных. Этот метод нашел применение в сельском хозяйстве. Для массовой работы с овцами бралась сыворотка крови жеребой кобылы (СЖК), содержащая гормон гипофиза; инъекция этой сыворотки за 2—5 дней до естественной овуляции увеличивала продукцию яиц почти вдвое, что выражалось в значительном учащении случаев многоплодия, как это, например, показывает диаграмма, обобщающая работу над отарой овец в одном из совхозов Средней Азии (рис. 20). Приплод благодаря методу Завадовского в среднем увеличивается на 50 и более ягнят на каждые 100 маток. В некоторых случаях вместо 1—2 ягнят в контрольной группе подопытные овцы приносили по 5 и 6 за раз. Методом Завадовского многоплодие, очевидно, достигается благодаря образованию РБ. Физиологический механизм, на котором основан метод, в основном ясен, хотя для ряда деталей вопроса требуются еще дополнительные исследования.
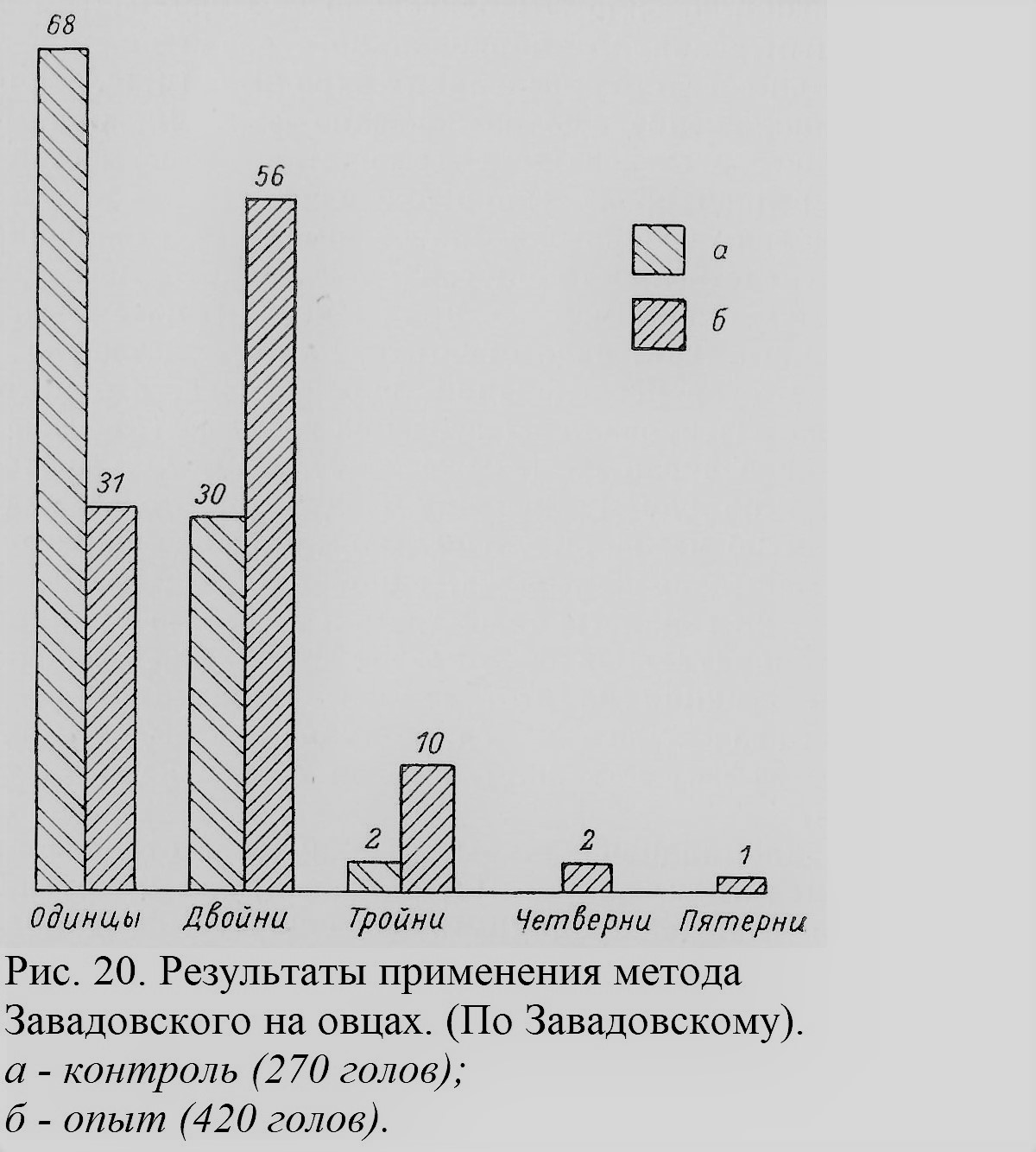
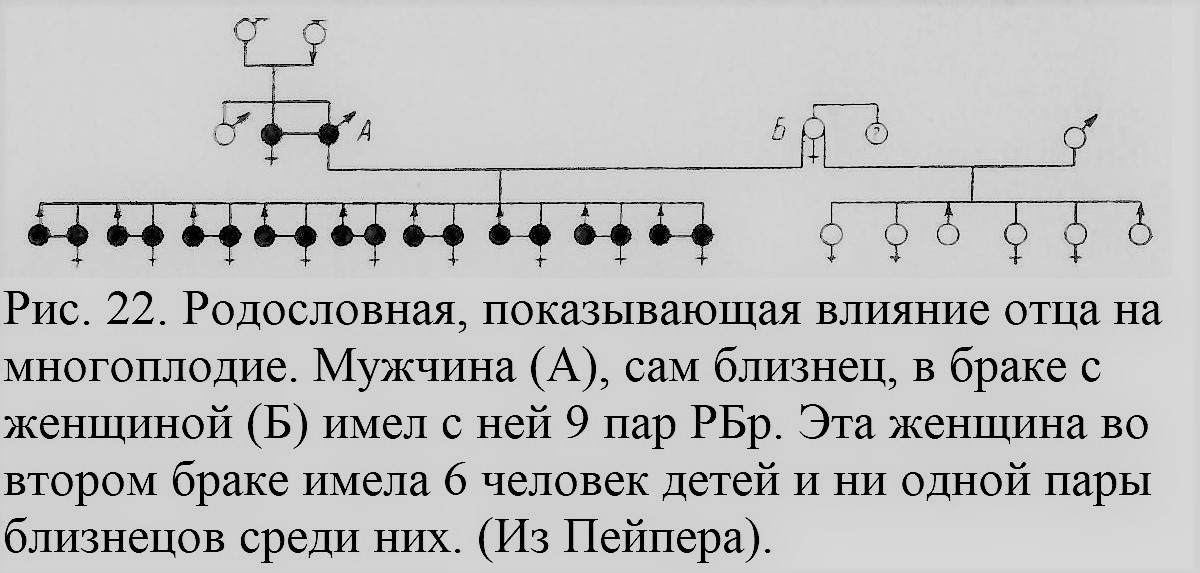
Некоторые авторы ищут возможных связен склонности к многоплодию с плодовитостью как таковой (Gedda, 1951). Само понятие плодовитости недостаточно ясно, и весь вопрос требует еще дальнейшего изучения. Конечно, условия, влияющие на одновременное созревание нескольких яиц, на их оплодотворение, дальнейшее развитие зиготы,[1] ее инплантацию [2]в стенку матки и т. д., имеют прямое отношение к РБ (Hammond, 1941). Но здесь мы этими вопросами заниматься не можем. Особым вопросом является возможная зависимость между возникновением ОБ и РБ. Мы его уже касались не раз. Есть предположение, что, допуская физиологическое различие и независимость процессов образования ОБ и РБ (вопреки гипотезе Дальберга и других), все же наличие РБ может стимулировать возникновение ОБ благодаря тем трудностям и неблагоприятным условиям эмбрионального развития, которые возникают при соседстве нескольких зародышей (McArthur, 1942). Это предположение еще не доказано фактами, и необходимы дальнейшие исследования возможных зависимостей между возникновением обоих типов близнецов. Рассмотрим теперь ряд фактов, имеющих отношение к проблеме наследственности многоплодия. Некоторые из них обнаруживаются, например, и при изучении материала опытов Завадовского. Так, надо отметить, как это видно из рис. 20, и общих итогов исследований Завадовского, что не все овцы одинаково реагировали на введение гормона гипофиза (СЖК): значительное число их (свыше 30%), несмотря на инъекцию, приносило по одному ягненку («одинцу»), тогда как другие приносили по нескольку ягнят. Помимо ряда моментов, которые подробнее не изучались, одной из причин такой разницы в реакции на гонадотропный гормон гипофиза является, вероятно, индивидуальное своеобразие отдельных особей. Детали различия реакции на этот гормон не изучались, поскольку интерес Завадовского и его сотрудников был направлен на массовый эффект инъекции в связи с сельскохозяйственными задачами. Изучение индивидуальных различий в отношении многоплодия отдельных животных, как овец, так и других, позволило бы глубже проникнуть в сущность физиологического механизма явлений многоплодия, и, вероятно, это станет задачей будущих исследований. У людей такие индивидуальные различия не раз отмечались, особенно в тех случаях, когда женщина проявляла исключительную способность к многоплодным родам. Еще Аристотель в своей «Истории животных» (кн. VII, гл. 4) пишет об одной женщине, родившей 20 детей, в каждые роды по 5 человек. Подобные сообщения сохранились в различных документах минувших веков; степень достоверности их порой сомнительна. Приведем лишь несколько примеров из недавнего прошлого. Так, одна уроженка Вены имела 11 родов, все многоплодные: 3 двойни, 6 троен и 2 четверни. Одна арабская женщина в Марокко имела 5 родов, произведя на свет 4 двойни и одну тройню, все одинакового пола. Описан случай, когда одна женщина 33 лет имела 44 ребенка, все рожденные в многоплодных родах — 13 двоен и 6 троен (Caullery, 1945). В ряде случаев многоплодные роды чередуются весьма по-разному с одиночными родами. Предрасположение данной женщины к многоплодным родам особенно ярко обнаруживается в тех случаях, когда она вступает в брак дважды или трижды и, несмотря на разных мужей, все же рожает близнецов. Примером может служить женщина, на рис. 21, обозначенная III, 3, в приводимой здесь родословной из Дэвенпорта (Davenport, 1919). В некоторых случаях установлено также влияние отца на многоплодие. В доказательство приводятся такие факты разной степени достоверности (Bell, 1933). В конце XVIII в., например, жил русский крестьянин Федор Васильев, имевший от первой жены 4 четверни, 7 троен и 16 двоен, а от второй жены 2 тройни и 6 двоен, всего 87 детей, из которых 84 остались в живых. Другой аналогичный случай — тоже русский крестьянин «Кирлов» (Kyrloff), живший в середине XVIII в. и от двух жен имевший 72 ребенка; все они явились на свет в многоплодных родах. Еще один мужчина, сам будучи разнояйцевым близнецом, имел 9 разнояйцевых пар близнецов от женщины, которая, выйдя замуж вторично, имела от второго мужа 6 детей-одиночек и ни одной пары близнецов (рис. 22; Peiper, 1923).

Природа влияния мужчины на многоплодие не ясна (McArthur, 1942). Кроме того, подобных фактов исключительного многоплодия мало, и они недостаточно изучены, так как нет сведений относительно не только самих этих лиц, но также их родственников разных степеней родства. Это важно потому, что такие случаи индивидуальной склонности к многоплодию оказываются не единичными в некоторых семьях (рис. 23), а в той или иной форме встречаются среди родственников родителей близнецов, как отцов, так и матерей. Примером может служить потомство некой Бершерс, имевшей дочерей — ОБ, которые, в свою очередь, в один день 1947 г. родили по паре РБ (рис. 24).


Такие факты вызывают предположение, что склонность к многоплодию наследственна (Gedda, 1951). В связи с этим еще со второй половины XIX в. был осуществлен ряд исследований семей, где встречаются близнецы, составлены родословные и статистически изучен состав таких семейств. В этих исследованиях было поднято несколько вопросов наследственности многоплодия. Из них важнейшие: идет ли склонность к многоплодию со стороны матери, отца или обоих родителей? Является ли наследственным рождение только одного из двух типов близнецов или обоих? Обусловлена ли наследственность РБ и ОБ одним общим фактором или разными? и т. д. Роль матери, которая сама является близнецом или имеет среди родни близнецов, как будто ясно выступает в различных родословных, как например в упомянутой родословной, описанной Дэвенпортом (рис. 21). Другие родословные, приводимые Дэвенпортом (Davenport, 1920, 1928), служат для демонстрации роли отца, имеющего близнецов среди своей родни. Такие родословные описаны и другими авторами (Meyer, 1932; Greulich, 1930, 1938, и др.). На основании изучения 200 родословных Дэвенпорт нашел, что влияние отца на рождение близнецов лишь несколько слабее материнского влияния. В семьях, производящих близнецов, среди братьев-сестер матери имелось 7.8% близнецов, среди таковых отца — 6.5%. Среди матерен близнецов 5.3% были в свою очередь близнецами, среди отцов близнецов — 2.9%. Что касается потомства самих близнецов, то у дочерей их было 22.7% близнецов, а у сыновей близнецов — 21.7%. Аналогичные расчеты приводили и другие авторы (Verschuer, 1932; Gedda, 1951, и др.). Приводим еще таблицу Гедды (Gedda, 1951), составленную на основании серий родословных без специального подбора таковых. В таком подборе упрекали других авторов, так как это повышало относительное количество близнецов в родне исходных в исследовании близнецов (пробатов). Гедда использовал сравнительно небольшой материал, почерпнутый из населения Рима (табл. 4).
 Из общего числа близнецов среди родни исходных в исследовании близнецов около половины происходит из рода матери (50.58%), около четверти — из рода отца (25.88%) и сравнительно немного в семьях,в которых близнецы имелись со стороны обоих родителей (15.29%). Выявляется малопонятное различие между РБ и ОБ: у РБ процент близнецов среди родни матери значительно больше, чем среди родни отца (56.81 к 20.45). Эта разница меньше у ОБ (43.90 к 31.70).
Для сравнения с цифрами этой таблицы Гедда приводит еще данные, полученные из другого пункта Италии (Джези), которые, несмотря на некоторые различия, в общем аналогичны вышеприведенным. На основании своих материалов Гедда считает, что многоплодие наследственно со стороны обоих родителей для обоих типов близнецов. Эту точку зрения уже высказывали другие исследователи (Curtius u. Verschuer, 1932; отчасти — Meyer, 1932, и др.), однако она далеко не всеми разделяется. Одни отрицают всякую наследственность такого рода (Lenz, 1933). Другие допускают вероятность только наследования склонности к разнояйцевым родам и по женской линии (Bonnevie a. Sverdrup, 1926), третьи, наконец, считают вопрос в целом еще недостаточно изученным и пока что открытым (Lotze, 1937; Caullery, 1945, и др.). На основании того материала, который в настоящее время известен относительно этого вопроса у человека, последняя точка зрения кажется наиболее приемлемой. И пока различные причины, влияющие на многоплодие, достаточно подробно и полно, не изучены, доказательство наследственности многоплодия останется плохо обоснованным.
Наследственность многоплодия у животных тоже еще недостаточно исследована. Есть, например, ряд данных относительно овец — между многоплодием матерей и дочерей существует будто бы явная связь. Имеются указания, что на плодовитость влияет также отец. Завадовский (1941) считает, что наследственная природа многоплодия не может подлежать сомнению, но вопрос требует дальнейшего изучения, так как детали закономерностей наследования не известны. Этот же вопрос изучался на коровах, но также еще недостаточно (W. Weber и др.).
Многоплодие в пределах вида стоит в связи с размерами тела разных рас, пород и линий. Отбор с целью выведения крупных форм или, наоборот, мелких животных того же вида обнаруживает эту зависимость многоплодия от величины тела. Например, велась селекция на домашних мышах одной популяции с целью получить две линии разной величины, также на кроликах (McArthur, 1942). Кролики мелкой расы (польской), самки которых весят 1508 г., имели в среднем 3.2 детеныша в помете, тогда как крупная раса (красная ново-зеландская), самки которой весят 5589 г., имела в среднем 10.2 детеныша в помете. Аналогичные данные есть для свиней, собак и т. д. По-видимому, и среди диких животных существуют подобные явления. В природе они осложняются еще зависимостью от климата, так как в северных широтах и горных местностях млекопитающие и птицы имеют более крупные размеры, чем те же виды в теплых районах (правило Бергмана).
Мак-Артур сопоставляет наблюдения над животными с некоторыми данными о человеке. Еще Чурилов (1878) сделал попытку показать, что многоплодие у человека увеличивается в соответствии с увеличением роста группы населения. Свои выводы он основывал на изучении некоторых европейских народов (итальянцев, французов и др.). Такой параллелизм между ростом и многоплодием не был обнаружен на американском и новозеландском населении (Apert, 1923). По расчетам Дальберга (Dahlberg, 1926), есть положительная корреляция между светлым цветом глаз (серый, голубой) некоторых европейских народов и рождением у них близнецов. Светлоглазыми являются северные племена, обычно более крупного роста. Среди других народов негры, например, отличающиеся значительным ростом, чаще имеют близнецов, чем японцы, — народ сравнительно малорослый, имеющий приблизительно одни близнецовые роды на 300 обычных. Однако все эти сопоставления роста с многоплодием еще не решают вопроса, чем вызвана такая зависимость, если она реально существует: меньшим числом многоплодных зачатий у людей с малым ростом или трудностью беременности двойней для малорослых, как думал Чурилов.
Этим мы затрагиваем вопрос о частоте рождения близнецов у разных рас, как наследственном признаке таковых. Этот вопрос еще слишком недостаточно изучен, чтобы можно было делать какие-нибудь обобщения, тем более, что многоплодие не всегда непосредственно зависит от размеров тела.
Вопрос о расовых свойствах современных народов, как известно, чрезвычайно осложнен многими обстоятельствами: смешением рас, различиями их расселения на земле, социально-экономическими условиями и т. д. И даже если можно считать равными географические и климатические условия для негроидов и европеоидов, живущих в США, то все же сравнение многоплодия этих двух групп, которое делают некоторые исследователи (Strandskov a. Edelen, 1946; Enders a. Stern, 1948, и др.), настолько обусловлено социально-экономическими моментами, что считать существующее различие в многоплодии расовым признаком нельзя. Чтобы это доказать, надо взять материал сравниваемых рас и групп населения, одинаковых в социально-экономическом отношении; этого американские авторы не делают.
Очевидно, социально-экономические факторы являются решающими в отношении несколько большего количества близнецов, в основном РБ, рождавшихся в деревне по сравнению с городом в первой четверти нашего века (Weinberg, 1902; Bonnevie, 1919, и др.). Это объясняется вообще большей рождаемостью в деревне, чем в городе; и, в частности, многодетностью немолодых матерей, у которых, как мы знаем, с возрастом увеличиваются шансы на рождение близнецов. Их сверстницы-горожанки, по-видимому, различными способами и по различным мотивам чаще уклоняются от деторождения, что, конечно, не может не отзываться на относительном уменьшении рождаемости близнецов.
Статистически также установлено, что среди незаконнорожденных детей близнецы встречаются несколько реже, чем среди законнорожденных (Weinberg, 1902; Dahlberg, 1926; Gedda, 1951, и др.). Дальберг это объясняет обычно более молодым возрастом матерей незаконных детей.
Вообще роль социально-экономических факторов в вопросе о многоплодии еще мало изучена и этим факторам уделяется недостаточное внимание.
Из общего числа близнецов среди родни исходных в исследовании близнецов около половины происходит из рода матери (50.58%), около четверти — из рода отца (25.88%) и сравнительно немного в семьях,в которых близнецы имелись со стороны обоих родителей (15.29%). Выявляется малопонятное различие между РБ и ОБ: у РБ процент близнецов среди родни матери значительно больше, чем среди родни отца (56.81 к 20.45). Эта разница меньше у ОБ (43.90 к 31.70).
Для сравнения с цифрами этой таблицы Гедда приводит еще данные, полученные из другого пункта Италии (Джези), которые, несмотря на некоторые различия, в общем аналогичны вышеприведенным. На основании своих материалов Гедда считает, что многоплодие наследственно со стороны обоих родителей для обоих типов близнецов. Эту точку зрения уже высказывали другие исследователи (Curtius u. Verschuer, 1932; отчасти — Meyer, 1932, и др.), однако она далеко не всеми разделяется. Одни отрицают всякую наследственность такого рода (Lenz, 1933). Другие допускают вероятность только наследования склонности к разнояйцевым родам и по женской линии (Bonnevie a. Sverdrup, 1926), третьи, наконец, считают вопрос в целом еще недостаточно изученным и пока что открытым (Lotze, 1937; Caullery, 1945, и др.). На основании того материала, который в настоящее время известен относительно этого вопроса у человека, последняя точка зрения кажется наиболее приемлемой. И пока различные причины, влияющие на многоплодие, достаточно подробно и полно, не изучены, доказательство наследственности многоплодия останется плохо обоснованным.
Наследственность многоплодия у животных тоже еще недостаточно исследована. Есть, например, ряд данных относительно овец — между многоплодием матерей и дочерей существует будто бы явная связь. Имеются указания, что на плодовитость влияет также отец. Завадовский (1941) считает, что наследственная природа многоплодия не может подлежать сомнению, но вопрос требует дальнейшего изучения, так как детали закономерностей наследования не известны. Этот же вопрос изучался на коровах, но также еще недостаточно (W. Weber и др.).
Многоплодие в пределах вида стоит в связи с размерами тела разных рас, пород и линий. Отбор с целью выведения крупных форм или, наоборот, мелких животных того же вида обнаруживает эту зависимость многоплодия от величины тела. Например, велась селекция на домашних мышах одной популяции с целью получить две линии разной величины, также на кроликах (McArthur, 1942). Кролики мелкой расы (польской), самки которых весят 1508 г., имели в среднем 3.2 детеныша в помете, тогда как крупная раса (красная ново-зеландская), самки которой весят 5589 г., имела в среднем 10.2 детеныша в помете. Аналогичные данные есть для свиней, собак и т. д. По-видимому, и среди диких животных существуют подобные явления. В природе они осложняются еще зависимостью от климата, так как в северных широтах и горных местностях млекопитающие и птицы имеют более крупные размеры, чем те же виды в теплых районах (правило Бергмана).
Мак-Артур сопоставляет наблюдения над животными с некоторыми данными о человеке. Еще Чурилов (1878) сделал попытку показать, что многоплодие у человека увеличивается в соответствии с увеличением роста группы населения. Свои выводы он основывал на изучении некоторых европейских народов (итальянцев, французов и др.). Такой параллелизм между ростом и многоплодием не был обнаружен на американском и новозеландском населении (Apert, 1923). По расчетам Дальберга (Dahlberg, 1926), есть положительная корреляция между светлым цветом глаз (серый, голубой) некоторых европейских народов и рождением у них близнецов. Светлоглазыми являются северные племена, обычно более крупного роста. Среди других народов негры, например, отличающиеся значительным ростом, чаще имеют близнецов, чем японцы, — народ сравнительно малорослый, имеющий приблизительно одни близнецовые роды на 300 обычных. Однако все эти сопоставления роста с многоплодием еще не решают вопроса, чем вызвана такая зависимость, если она реально существует: меньшим числом многоплодных зачатий у людей с малым ростом или трудностью беременности двойней для малорослых, как думал Чурилов.
Этим мы затрагиваем вопрос о частоте рождения близнецов у разных рас, как наследственном признаке таковых. Этот вопрос еще слишком недостаточно изучен, чтобы можно было делать какие-нибудь обобщения, тем более, что многоплодие не всегда непосредственно зависит от размеров тела.
Вопрос о расовых свойствах современных народов, как известно, чрезвычайно осложнен многими обстоятельствами: смешением рас, различиями их расселения на земле, социально-экономическими условиями и т. д. И даже если можно считать равными географические и климатические условия для негроидов и европеоидов, живущих в США, то все же сравнение многоплодия этих двух групп, которое делают некоторые исследователи (Strandskov a. Edelen, 1946; Enders a. Stern, 1948, и др.), настолько обусловлено социально-экономическими моментами, что считать существующее различие в многоплодии расовым признаком нельзя. Чтобы это доказать, надо взять материал сравниваемых рас и групп населения, одинаковых в социально-экономическом отношении; этого американские авторы не делают.
Очевидно, социально-экономические факторы являются решающими в отношении несколько большего количества близнецов, в основном РБ, рождавшихся в деревне по сравнению с городом в первой четверти нашего века (Weinberg, 1902; Bonnevie, 1919, и др.). Это объясняется вообще большей рождаемостью в деревне, чем в городе; и, в частности, многодетностью немолодых матерей, у которых, как мы знаем, с возрастом увеличиваются шансы на рождение близнецов. Их сверстницы-горожанки, по-видимому, различными способами и по различным мотивам чаще уклоняются от деторождения, что, конечно, не может не отзываться на относительном уменьшении рождаемости близнецов.
Статистически также установлено, что среди незаконнорожденных детей близнецы встречаются несколько реже, чем среди законнорожденных (Weinberg, 1902; Dahlberg, 1926; Gedda, 1951, и др.). Дальберг это объясняет обычно более молодым возрастом матерей незаконных детей.
Вообще роль социально-экономических факторов в вопросе о многоплодии еще мало изучена и этим факторам уделяется недостаточное внимание.
Глава четвертая
ВОЗНИКНОВЕНИЕ БЛИЗНЕЦОВ
Название двух основных типов близнецов — ОБ и РБ — уже говорит о разном их возникновении, которое и создает основное различие этих типов. Теперь нам надо остановиться на нем подробнее. Можно сказать, что если РБ являются результатом полиовуляции, т. е. образования нескольких яиц, приблизительно одновременно оплодотворенных, то ОБ являются результатом полиэмбрионии, т. е. образования нескольких зародышей из одного оплодотворенного яйца (зиготы). Поэтому РБ некоторые ученые склонны рассматривать как одновременно родившихся нескольких обыкновенных братьев и сестер, по существу ничем не отличающихся от них. ОБ являются результатом коренного изменения обычного для большинства животных развития зародыша, превращения его в несколько индивидов, вместо превращения его в одного. Процесс возникновения ОБ сложнее такового РБ, протекает разнообразно и далеко еще не вполне выяснен. Мы знаем, что у известных животных образование ОБ является закономерным явлением, как например у некоторых паразитических перепончатокрылых или у армадилов; это явление Паттерсон (Patterson, 1927) назвал «специфической полиэмбрионией, в отличие от «спорадической», изредка наблюдающейся у животных, обычно имеющих одного детеныша (лошадь, корова и т. д.). Наконец, можно выделить, как делает Паттерсон, «экспериментальную» полиэмбрионию, вызванную искусственным вмешательством, как например перетяжка зиготы тритона и т. п. (см. главу восьмую «млекопитающие»). Прежде всего, чтобы приблизиться к пониманию механизма возникновения ОБ, следует остановиться на том, на какой стадии эмбрионального развития происходит разделение сначала единого зародыша на два или несколько индивидов. Из приведенного далее материала видно, что в эксперименте это может происходить начиная со стадии первых двух бластомеров; так бывает у морских ежей, тритонов и даже кроликов. Обособление отдельных бластомеров или групп бластомеров происходит и у перепончатокрылых, и у других животных. Близнецы, как известно, возникают и на более поздних стадиях, например у рыб, птиц и других животных во время образования бластодермы.[3] Наконец, близнецы возникают и на еще более поздних стадиях - перед гаструляцией[4] и в начале ее: в стадии бластоцисты[5] у млекопитающих (см. главу восьмую «млекопитающие»), также, вероятно, и у человека, до появления первичной полоски, как это описано у армадилов и как это можно предположить и у человека на основании известного препарата Стритера. На стадии ранней гаструляции ОБ возникают также у морских звезд, рыб, птиц и других животных. Но позже, на стадии поздней гаструляции и после неё, по-видимому, близнецы уже не возникают. Однако удвоение отдельных участков тела возможно иногда и на более поздних стадиях развития, например при развитии конечностей у тритонов. Чем вызывается раздвоение дробящейся зиготы или яйца на ранней стадии эмбриогенеза? На этот вопрос, вероятно, нельзя дать один общий ответ. У разных животных, при разных обстоятельствах, на различных стадиях могут быть, надо думать, разные причины раздвоения или разделения на большее число обособленных частей. Чисто механические причины могут вызвать, например, раздвоение зиготы морского ежа на стадии двух бластомеров. Аналогичный эффект механической изоляции двух половин зиготы или бластулы тритона вызывает перетяжка волосом и другие подобные экспериментальные вмешательства. Однако, например, распад зародыша перепончатокрылого на отдельные группы бластомеров является, вероятно, результатом каких-то физиологических процессов, пока не изученных, приводящих к физиологической изоляции этих групп бластомеров, т. е. в известной мере к физиологической независимости таких групп клеток друг от друга. Физиологической изоляцией объясняют и различные явления раздвоения сначала единого зачатка на стадии до гаструляции и в начале нее. Единство развития зародыша зависит, согласно теории физиологического градиента, от возникновения в развивающемся яйце участка с максимальной активностью обмена, играющего ведущую роль в развитии. Такой участок оказывается также наиболее уязвимым вредящими факторами, как холод, ядовитые вещества и т. и. После временного действия таких факторов и восстановления нормальных условий могут возникнуть новые градиенты в числе двух, трех и больше, и гаструляция может произойти в более чем одном месте (рис. 25; ср. Барт, 1951).

Такие явления описаны у морских звезд, птиц и других животных (см. главу восьмую «Беспозвоночные»). Повреждающими факторами, временно останавливающими развитие, являются недостаток кислорода, холод и другие внешние агенты, встречающиеся в естественных условиях или искусственно применяемые в эксперименте, например ультрафиолетовое облучение или ядовитые вещества. Такие факты получены в опытах Стоккарда с рыбами, в опытах Ньюмена с морскими звездами. Эти внешние факторы, вызывающие возникновение ОБ, однако, действуют в разной степени эффективно в зависимости от вида животных; так, например, у одного вида лягушек (Rana pipiens) холод, недостаток кислорода и облучение вовсе не могли вызвать возникновения близнецов (Schwind, 1942), как у рыб в вышеупомянутых опытах. Одна из «внутренних» причин возникновения ОБ — это перезревание яйца. Как показал Витши (Witschi, 1934), яйца лягушки, оплодотворенные через 3—5 дней после овуляции, проявляют заметную способность к образованию ОБ. Это объясняется тем, что «физиологические корреляции», под влиянием которых развивается нормальный эмбрион, оказываются ослабленными «старостью» яиц. Далее, в связи с этим возникают физиологически обособленные участки, играющие роль самостоятельных «организаторов», что ведет к образованию ОБ и аномалий развития. Со стороны именно таких «внутренних», физиологических зависимостей процесс возникновения ОБ еще очень мало исследован. Для понимания возникновения человеческих ОБ большой интерес представляет процесс образования ОБ у броненосцев-армадилов, единственных млекопитающих, у которых это явление изучено полнее, чем у других. Раздвоение у этих животных происходит еще до образования первичной полоски, т. е. на сравнительно ранней стадии эмбриогенеза. Самый же способ раздвоения, разный даже у двух видов армадилов, вероятно, настолько специфичен у каждого из них, что предполагать подобный способ у других млекопитающих вряд ли допустимо. Физиологический механизм раздвоения и у армадилов еще мало понятен, тем более у других млекопитающих. Что касается стадии эмбриогенеза, когда происходит возникновение ОБ у человека, то, вероятно, она не всегда одна и та же, а несколько колеблется, то приходясь на стадии бластоцисты (рис. 26) и даже раньше, то несколько позже, перед развитием первичной полоски, но после образования амниона[6], как у армадила. Об этом можно судить на основании оболочек у ОБ и по строению соединенных близнецов, которые в ряде случаев считаются результатом позднего, не закончившегося раздвоения зародыша (Newman, 1931).

Интересно, что раздвоение ранней стадии зародыша у человека может происходить несколько раз повторно — иначе нельзя объяснить возникновение однояйцевых четверен и пятерен. Так, например, для генезиса известной пятерни Дионн (см. главу девятую) предлагается следующая гипотетическая схема (рис. 27), основанная на некоторых чертах сходства и различия между этими близнецами (Newman, 194(4). У этой пятерни было всего три амниона, в которых находились один, два и два зародыша, что и соответствует приведенной схеме.

Возникновение РБ объяснить легче, чем возникновение ОБ. Основное явление состоит здесь, как уже говорилось, в более или менее одновременном оплодотворении двух или более яиц. Различие может быть только в способах образования этих яиц. Самый обычный — это одновременное созревание нескольких яиц в обоих яичниках животного, каждое яйцо в своем фолликуле. Как известно, лопнувший при овуляции фолликул превращается в желтое тело, и по числу их в яичниках можно судить о числе яиц, поступивших в яйцеводы. Обычно число РБ соответствует числу желтых тел, но может быть и меньше числа последних, если часть яиц погибла, не превратившись в зародышей. Существует еще другая возможность, вообще более редкая, это образование двух или более яиц в одном фолликуле, с образованием одного желтого тела. Подобные случаи известны у животных и у человека. Так, например, описаны яичники одной 18-летней женщины, у которой в фолликулах имелось по нескольку (от 2 до 12 ооцитов); некоторые из них были крупнее, другие мельче и, очевидно, отставали в развитии. В другом случае у убитой 25-летней женщины при вскрытии были обнаружены близнецы мужского пола в двух хорионах при наличии одного желтого тела в одном из яичников (Caullery, 1945); в другом сходном случае близнецы были разного пола (Komai a. Fukuoka, 1937). Ряд подобных случаев описан многими авторами (ср. Miettinen, 1954). При полиовуляции возможны два варианта оплодотворения: яйца оплодотворяются спермой одного самца или же разных (суперфекундация). У животных, как кошки или собаки, последняя возможность, по-видимому, не так уж редко наблюдается. Предполагается она в некоторых случаях и у человека, например при рождении белой женщиной пары РБ, состоящей из черного и белого младенца (Gedda, 1951; Miettinen, 1954). Однако такие случаи очень редки, недостаточно хорошо изучены, и потому интерпретация их еще спорна. Наконец, наблюдалась еще третья возможность одновременного образования двух яиц, еще более редкая — это образование второго редукционного тельца,[7] столь большого размера, что оно могло играть роль второго яйца и оплодотворяться другим спермием. Иначе говоря, ооцит во время деления созревания превращается в две почти равновеликие клетки, «двойное яйцо», вместо того чтобы, как обычно, образовывать крупное яйцо и маленькое редукционное тельце. Такое явление наблюдалось у морских ежей (Lindahl, 1937), кольчатых червей (Tubifex) и даже мышей (Gates, 1946). Высказывалось предположение, пока не доказанное, что оно возможно и у людей (Lehmann u. Huber, 1944). На это явление опиралась также гипотеза о том, что возникновение РБ п ОБ имеет общую основу — наследственную «тенденцию к делению» (Dahlberg, 1926—1952; Verschuer, 1932; Curtius u. Verschuer, 1932, и др.). Эта «тенденция» выражается при созревании яйца в образовании двух яиц вместо одного яйца и редукционного тельца, т. е. в диовуляции и образовании РБ, а позже, в другом случае — в раздвоении эмбриона, в диэмбрионии, т. е. образовании ОБ. Эту гипотезу до сих пор не удалось доказать, как признается один из авторов ее — Дальберг (Dahlberg, 1952). О гормональном механизме, вызывающем обычную полиовуляцию, уже говорилось выше. Этим несомненно сделан реальный шаг к пониманию физиологической основы возникновения РБ. В литературе встречаются еще некоторые гипотезы, ныне устаревшие, для объяснения возникновения близнецов как бы промежуточных между двумя основными типами: это оплодотворение двуядерного яйца двумя спермиями (Тур, 1904б), что достоверно не было доказано, и если и возможно, то очень редко (Gates, 1946), или же просто оплодотворение одного яйца несколькими спермиями, что в настоящее время признано ошибочным предположением (Caullery, 1945), так же как оплодотворение двухголовым спермием и т. п. (Schwalbe, 1907).

Нам уже неоднократно приходилось попутно говорить об оболочках близнецов. Теперь надо остановиться на них несколько подробнее, тем более, что по ним можно отчасти судить о стадии развития эмбриона, на которой он раздвоился, образуя ОБ, как об этом уже упоминалось выше. Кроме того, оболочки долго служили в глазах акушеров решающим критерием в вопросе, является ли данная пара близнецов ОБ или РБ, так как считалось, что РБ всегда имеют разные хорионы, а ОБ — всегда имеют общий хорион (рис. 28). Однако это обобщение, основанное, казалось бы, на огромном опыте родильных учреждений разных стран, после тщательной проверки, в связи с развитием изучения близнецов, оказалось неверным. Выяснилось, что могут быть ОБ с разными хорионами «дихориальные», и их около 20% среди всех ОБ (Rife, 1952а). Было поставлено специальное исследование близнецов и их оболочек (Gurtius, 1930). Для того чтобы избежать предвзятого мнения о типе близнецов, одна группа ученых занималась только изучением оболочек и по ним решала вопрос о типе близнецов, другая изучала только самих близнецов и решала тот же вопрос на основании внутрипарного сходства близнецов, т. е. сходства между близнецами каждой пары, по методу, о котором речь подробно будет дальше (глава четырнадцатая). Для этого близнецы обследовались не только после рождения, но и позже, так как не все нужные для сравнения признаки обнаруживаются у новорожденных. Результаты для близнецов внутрипарно одного пола сведены в табл. 5 (Lotze, 1937).
 Мы видим, что из 100 дихориальных пар 24 были ОБ; все монохориальные — тоже ОБ. Большинство ОБ имело при одном хорионе два амниона. Лишь очень немногие пары (3) имели один общий и хорион, и амнион. Все РБ имели разные хорионы и, разумеется, амнионы. Таким образом, можно различать три рода ОБ но оболочкам. Предположительно можно эти типы объяснить тем, что раздвоение произошло у них на разных стадиях развития (Verschuer, 1932, 1939; Caullery, 1945, и др.). Известно, что сначала закладывается хорион, затем амнион и собственно зародыш. Поэтому можно предположить, что те ОБ, которые имеют разные хорионы и разные амнионы, как РБ, возникли на самых ранних стадиях развития, может быть, из первых бластомеров; имплантация в стенку матки, как и дальнейшее развитие, происходила для каждой половины первоначального яйца самостоятельно. Далее, монохориальные ОБ, но с двумя амнионами возникли, вероятно, из одного имплантировавшегося яйца, но позже зародышевый узелок раздвоился, и в одном хорионе возникло два эмбриона с двумя амнионами, как показано на вышеприведенной схеме (рис. 26, А). Наконец, монохориальные и моноамниотические ОБ — это, по-видимому, такие, которые являются самыми поздними по времени возникновения, после образования амниотической полости, приблизительно на той стадии, на которой возникают близнецы у армадила. Так как соединенные близнецы обычно имеют один амнион, то и их склонны считать продуктами позднего раздвоения эмбриона, также с разными сдвигами в ту или другую сторону. Как известно, соединенные близнецы редки. Связь между оболочками и двойными уродствами заметно выступает из следующих данных (Steiner, 1935): среди 24 пар ОБ дихориальных был один случай уродства, среди 32 пар ОБ монохориальных с двумя амнионами — 3 случая, а из трех пар ОБ с одним хорионом и одним амнионом — 2 случая уродств.
Основная масса ОБ, вероятно, относится к типу монохориальному, но диамниотическому, следовательно, возникающему уже после имплантации яйца в стенку матки. Дальнейшее изучение этого вопроса должно дополнить и уточнить известное в настоящее время.
При близком расположении двух зародышей в матке, особенно же в фаллопиевых трубах, оболочки могут слипаться, частично рассасываться, и полости двух хорионов или амнионов сообщаться. Это будет вторичное образование общей оболочки у разных зародышей, — иногда РБ, иногда даже разного пола (Агеу, 1922а). Такие явления наблюдаются сравнительно редко (Verschuer, 1932, и др.).
Что касается плаценты, то она у РБ чаще разная, но может срастаться в зависимости от степени близости имплантации яиц. У ОБ она чаще общая или сросшаяся; лишь редко она может быть разной, как у РБ; это бывает в некоторых случаях ОБ первого типа из только что рассмотренных — рано образовавшихся. Неодинаковое использование общей плаценты, анастомозы сосудов, снабжающих кровью обоих компонентов и т. п., — все это очень важные, иногда роковые для близнецов последствия общности плаценты. Подробнее об этом речь будет ниже.
Взаимоотношение между оболочками и разными типами близнецов, только что рассмотренное, может быть резюмировано в табл. 6, заимствованной из книги Лотце (Lotze, 1937) с незначительными дополнениями и изменениями.
Мы видим, что из 100 дихориальных пар 24 были ОБ; все монохориальные — тоже ОБ. Большинство ОБ имело при одном хорионе два амниона. Лишь очень немногие пары (3) имели один общий и хорион, и амнион. Все РБ имели разные хорионы и, разумеется, амнионы. Таким образом, можно различать три рода ОБ но оболочкам. Предположительно можно эти типы объяснить тем, что раздвоение произошло у них на разных стадиях развития (Verschuer, 1932, 1939; Caullery, 1945, и др.). Известно, что сначала закладывается хорион, затем амнион и собственно зародыш. Поэтому можно предположить, что те ОБ, которые имеют разные хорионы и разные амнионы, как РБ, возникли на самых ранних стадиях развития, может быть, из первых бластомеров; имплантация в стенку матки, как и дальнейшее развитие, происходила для каждой половины первоначального яйца самостоятельно. Далее, монохориальные ОБ, но с двумя амнионами возникли, вероятно, из одного имплантировавшегося яйца, но позже зародышевый узелок раздвоился, и в одном хорионе возникло два эмбриона с двумя амнионами, как показано на вышеприведенной схеме (рис. 26, А). Наконец, монохориальные и моноамниотические ОБ — это, по-видимому, такие, которые являются самыми поздними по времени возникновения, после образования амниотической полости, приблизительно на той стадии, на которой возникают близнецы у армадила. Так как соединенные близнецы обычно имеют один амнион, то и их склонны считать продуктами позднего раздвоения эмбриона, также с разными сдвигами в ту или другую сторону. Как известно, соединенные близнецы редки. Связь между оболочками и двойными уродствами заметно выступает из следующих данных (Steiner, 1935): среди 24 пар ОБ дихориальных был один случай уродства, среди 32 пар ОБ монохориальных с двумя амнионами — 3 случая, а из трех пар ОБ с одним хорионом и одним амнионом — 2 случая уродств.
Основная масса ОБ, вероятно, относится к типу монохориальному, но диамниотическому, следовательно, возникающему уже после имплантации яйца в стенку матки. Дальнейшее изучение этого вопроса должно дополнить и уточнить известное в настоящее время.
При близком расположении двух зародышей в матке, особенно же в фаллопиевых трубах, оболочки могут слипаться, частично рассасываться, и полости двух хорионов или амнионов сообщаться. Это будет вторичное образование общей оболочки у разных зародышей, — иногда РБ, иногда даже разного пола (Агеу, 1922а). Такие явления наблюдаются сравнительно редко (Verschuer, 1932, и др.).
Что касается плаценты, то она у РБ чаще разная, но может срастаться в зависимости от степени близости имплантации яиц. У ОБ она чаще общая или сросшаяся; лишь редко она может быть разной, как у РБ; это бывает в некоторых случаях ОБ первого типа из только что рассмотренных — рано образовавшихся. Неодинаковое использование общей плаценты, анастомозы сосудов, снабжающих кровью обоих компонентов и т. п., — все это очень важные, иногда роковые для близнецов последствия общности плаценты. Подробнее об этом речь будет ниже.
Взаимоотношение между оболочками и разными типами близнецов, только что рассмотренное, может быть резюмировано в табл. 6, заимствованной из книги Лотце (Lotze, 1937) с незначительными дополнениями и изменениями.
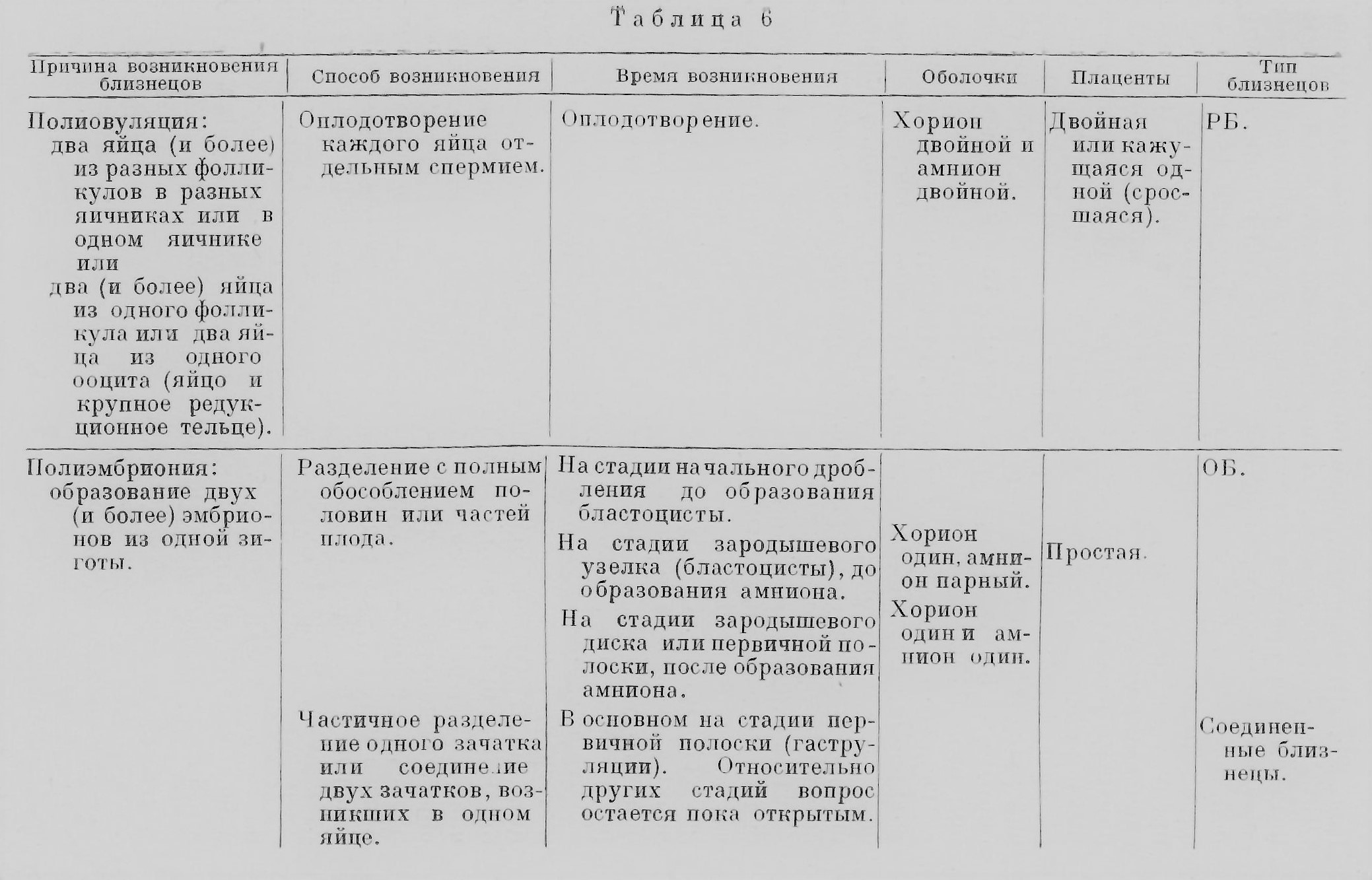
Глава пятая
ВОПРОСЫ УТРОБНОЙ ЖИЗНИ БЛИЗНЕЦОВ
Мы не будем останавливаться здесь на вопросах утробного развития близнецов различных животных, в норме имеющих несколько детенышей, являющихся, как мы знаем, обычно РБ. Для наших дальнейших целей имеет значение исследование условий утробной жизни близнецов у животных, в норме рождающих одного детеныша. Вопросы, сюда относящиеся, еще сравнительно мало изучены у таких животных, как корова или лошадь, отчасти ввиду большой редкости у них близнецов, особенно ОБ. На человеке, который нас более всего интересует, эти вопросы выяснены гораздо лучше. Поэтому мы и остановимся главным образом на вопросах утробной жизни человеческих близнецов. Матка женщины, как известно, приспособлена к развитию одного плода. Одновременное развитие двух и более плодов легко создает ненормальные отношения как между матерью и плодами, так и между плодами. Первая группа взаимоотношений, относящаяся преимущественно к области акушерства и гинекологии, меньше затрагивает интересующие нас вопросы, и потому мы здесь на них останавливаться не будем, хотя влияния материнского организма на плод могут быть очень многообразны и значительны и в этой связи существенны и для близнецов (ср. Bass, 1952; Grebe, 1956, и др.). Взаимоотношения между близнецами имеют очень важное значение для их развития и судьбы. Вопросы, возникающие из этих взаимоотношений, и составят содержание ближайших страниц. Близнецы могут располагаться в матке различно. Чаще всего они помещаются продольно (в 85—90% случаев), причем головы могут быть обращены или в одну сторону, или в противоположные (рис. 28). Поперечное положение одного из близнецов встречается в 10—15% случаев, поперечное положение обоих — очень редко, около 1 % (Verschuer, 1932; Gedda, 1951, и др.). От положения в матке в значительной мере зависит судьба близнецов. При невыгодном положении одного из близнецов по сравнению с другим, более удачливым, последний может обогнать в развитии своего партнера и, тесня его, довести до истощения и гибели. Это может произойти, в зависимости от многообразных конкретных условий, на разных стадиях эмбриональной жизни близнецов. В большинстве случаев РБ, чаще имеющие каждый свою плаценту и свои оболочки, меньше мешают друг другу в развитии, чем ОБ. Однако в некоторых случаях и у РБ, при наличии разных плацент, отставший в развитии близнец оказывается погибшим в зародышевом состоянии, обезвоженным и сжатым выжившим плодом, а потому выглядит как бы сделанным из бумаги — fetus papiraceus (рис.29).
Близкое расположение и соприкосновение плацент РБ иногда приводит к анастомозам кровеносных сосудов и гормональному влиянию через кровь одного плода на другой, как это известно у фри-мартинов (см. главу восьмую «копытные»); последние найдены не только у телят, но и у других млекопитающих, в том числе у обезьян и будто бы также и у человека (Gedda, 1951). Кроме того, смешение крови пары РБ таким путем, независимо от их пола, приводит иногда к возникновению своеобразных «химер» по кровяным группам или к «мозаицизму» эритроцитов. Это явление было описано у коров и овец (Owen, 1945; Anderson et al.,1951; Stormont et al., 1953), а недавно и у человека, пока только один случай (Dunsford et aI., 1953), касающийся женщины, у которой кровь давала реакцию на две группы: А и 0. У нее было около 61% эритроцитов группы 0. Эта женщина имела брата-близнеца, умершего в возрасте 3 месяцев. Сама она не была фри-мартином, так как имела вид нормальной женщины и была матерью одного ребенка. Возникновение «мозаицизма» эритроцитов до сих пор мало понятно. Существующая гипотеза о переходе кроветворных эмбриональных клеток из одного зародыша в другой благодаря анастомозам сосудов вызывает серьезные сомнения. Анастомозы сосудов играют очень большую роль в ряде случаев у ОБ, имеющих одну общую плаценту. Благодаря анастомозам между близнецами устанавливается своеобразный круг кровообращения, «третий круг» по Шатцу (Schatz, 1882, 1884—1910), подробно изучавшему его. Этим нарушается нормальное кровоснабжение эмбрионов, в результате чего могут возникнуть различные аномалии развития и гибель одного или обоих близнецов. Изучение кровеносной системы плаценты ОБ специальными методами (инъекция сосудов, рентгенограммы и др.) позволяет с большими подробностями установить картину анастомозов сосудов и распространение сосудов в плаценте (Gedda, 1951). Общая плацента далеко не всегда поровну используется обоими близнецами. Нередко наблюдается явно выраженная асимметрия — один из близнецов использует большую часть плаценты, чем другой (рис. 30).

Это уже не может не отразиться на развитии близнецов в связи с разницей в питании. Но, кроме того, в районе соприкосновения сосудов того и другого близнеца образуются анастомозы, которые тоже бывают далеко не всегда пропорциональными для артерий и вен. В результате может оказаться, что обмен крови через анастомозы между близнецами будет непропорционален: приток по артериям от близнеца I к близнецу II окажется больше, чем отток по венам обратно к I. Такая «динамическая асимметрия» в «третьем круге» ведет к нарастающему избытку крови в одном из близнецов и, соответственно, к недостатку крови в другом. Первый, имеющий избыток крови, развивается и растет лучше, чем второй. Однако этот избыток приносит и свой вред — гипертрофию сердца и печени, повышенное кровяное давление, утолщение стенок сосудов, избыток мочи и амниотической жидкости и т. д. Резко выраженный избыток крови приводит такого близнеца к гибели еще в эмбриональном состоянии. При более слабой степени этой асимметрии «третьего круга» такой близнец уже после рождения может иметь различные патологические изменения от избытка крови во время утробной жизни. Еще больше может пострадать второй близнец от недостатка крови. Он будет отставать в развитии в разной степени, в зависимости от степени «динамической асимметрии». У него кровяное давление будет понижено, появится недостаток мочи и т. д. Вместе с тем происходит и расстройство питания, что приводит отставшего близнеца к смерти. Вероятно, что смерть одного из ОБ, именно второго, с недостатком кровоснабжения, нередко вызвана именно асимметрией плацентарного «третьего круга» кровообращения (Newman, 1923; Wenner, 1947; Gedda, 1951, и др.). Одним из следствий описанной асимметрии является расстройство сердечной деятельности обездоленного близнеца и постепенная атрофия сердца, кончающаяся утратой сердца — частичной (hemiacardia) или полной (holoacardia). Различают несколько типов эмбрионов-близнецов, лишенных сердца, например форму, лишенную туловища (acardius acormus) (рис. 31), но имеющего голову или, наоборот, лишенного головы, с туловищем и недоразвитыми конечностями (acardius acephalus) (рис. 32). Лишенные сердца, эти зародыши могут лишь частично развиваться за счет кровоснабжения из «третьего круга» благодаря сердечной деятельности второго близнеца. Предельной формой недоразвития лишенного сердца близнецаявляется вовсе утративший внешние признаки человеческого тела неопределенного вида ком тканей (acardius amorphus), напоминающий опухоль (рис. 33) и, в сущности, аналогичный в известной мере «паразиту» при «автозите»[8] в системе асимметричных соединенных близнецов (см. глава шестая).

Партнер такого аморфного близнеца, по аналогии с автозитом, может быть относительно нормальным ребенком, которого при рождении легко счесть за одиночного новорожденного, поскольку его бесформенного близнеца трудно принять за человека. В результате подобных случаев происходит недоучет близнецовых родов в статистике. Некоторые считают, что только около половины близнецовых беременностей констатируются именно как близнецовые (Caullery, 1945). Рассмотренные отношения внутри пары ОБ в связи с развитием асимметрии плацентарного, «третьего круга» кровообращения являются лишь кратко изложенным примером весьма сложных и очень многообразных отношений между близнецами пары ОБ в силу их тесного соседства не только из-за образования «третьего круга», который вообще возникает у меньшинства ОБ. При удачном стечении обстоятельств оба партнера могут стать равноценными субъектами. Это оптимальный исход. Всевозможные уклонения от такого исхода, в разной форме и разной степени, дают картину различных аномалий, имеющих в пессимальном пределе дегенерацию и гибель одного, а иногда и обоих близнецов. Утробные условия жизни могут глубоко запечатлеваться в строении и функциях близнецов. Рождающаяся пара ОБ может иметь ряд внутрипарных различий, начиная от очень заметных, как разница в форме черепа (рис. 34), в весе, росте, общей жизнеспособности (рис. 35) и других свойств, до мелких различий, порой нелегко наблюдаемых. Эти различия могут на многие годы в разной мере проявляться и в дальнейшем, в жизни после рождения, и запечатлеться необратимо, на всю жизнь, в виде внутрипарной разницы не только роста или других физических особенностей, но и поведения двух членов пары ОБ, их характера, т. е. «склада» их высшей нервной деятельности (Канаев, 1954).

 Интересно отметить, что внутрипарные различия у ОБ оказываются в утробной жизни и при рождении в общем больше, чем у РБ. Это объясняется более тесным взаимодействием ОБ, чем РБ. В дальнейшей жизни такие различия ОБ, например роста и веса, легче сглаживаются, чем соответственные различия РБ (Lotze, 1937).
Все сказанное имеет большое методологическое значение при использовании «близнецового метода» (см. главу пятнадцатую), когда путем внутрипарного сравнения ОБ стремятся выяснить значение каких-нибудь факторов среды после рождения, т. е. в постнатальной жизни. Без достаточно точного и полного учета различий, возникших под воздействием условий утробной жизни, указанное сравнение может привести к ошибочным выводам. Используемые в исследованиях близнецы, дети и взрослые, часто оказываются неизученными в отношении различий, вызванных факторами утробной жизни, и это затрудняет или опорочивает соответствующие исследования в постнатальный период. Зависимость многих постнатальных внутрипарных различий от условий жизни до рождения — пренатальных — область, еще мало изученная, однако необходимая и очень важная для успешного развития так называемого «близнецового метода» генетики (Price, 1950; Grebe, 1952; Rife, 1952а, и др.).
Трудности утробной жизни близнецов проявляются в большем числе гибнущих в этот период зародышей, чем при обычных одиночных беременностях, выражаясь в преждевременных родах, выкидышах, мертворожденных и умирающих вскоре после рождения близнецов.
Гибель близнецов в утробный период трудно поддается учету. Можно предполагать, что более 25 % возникающих близнецов гибнет до рождения (Lotze, 1937). ОБ умирают относительно чаще, чем РБ, что объясняется большими трудностями утробной жизни первых. Аборты плода, одного или обоих, при близнецовых беременностях наблюдаются в несколько раз чаще, чем при обычных беременностях, например по американским данным в 5 раз (Hirst, 1940), по другим — в 2—3 раза (Gedda, 1951); эти цифры зависят от материала и способа учета абортов.
Преждевременные роды близнецов составляют значительный процент, но далеко не одинаковый для различных стран и мест (Gedda, 1951). В качестве примера могут служить данные французских исследователей (Gernez et Omez, 1938), полученные на 226 парах близнецов.
Интересно отметить, что внутрипарные различия у ОБ оказываются в утробной жизни и при рождении в общем больше, чем у РБ. Это объясняется более тесным взаимодействием ОБ, чем РБ. В дальнейшей жизни такие различия ОБ, например роста и веса, легче сглаживаются, чем соответственные различия РБ (Lotze, 1937).
Все сказанное имеет большое методологическое значение при использовании «близнецового метода» (см. главу пятнадцатую), когда путем внутрипарного сравнения ОБ стремятся выяснить значение каких-нибудь факторов среды после рождения, т. е. в постнатальной жизни. Без достаточно точного и полного учета различий, возникших под воздействием условий утробной жизни, указанное сравнение может привести к ошибочным выводам. Используемые в исследованиях близнецы, дети и взрослые, часто оказываются неизученными в отношении различий, вызванных факторами утробной жизни, и это затрудняет или опорочивает соответствующие исследования в постнатальный период. Зависимость многих постнатальных внутрипарных различий от условий жизни до рождения — пренатальных — область, еще мало изученная, однако необходимая и очень важная для успешного развития так называемого «близнецового метода» генетики (Price, 1950; Grebe, 1952; Rife, 1952а, и др.).
Трудности утробной жизни близнецов проявляются в большем числе гибнущих в этот период зародышей, чем при обычных одиночных беременностях, выражаясь в преждевременных родах, выкидышах, мертворожденных и умирающих вскоре после рождения близнецов.
Гибель близнецов в утробный период трудно поддается учету. Можно предполагать, что более 25 % возникающих близнецов гибнет до рождения (Lotze, 1937). ОБ умирают относительно чаще, чем РБ, что объясняется большими трудностями утробной жизни первых. Аборты плода, одного или обоих, при близнецовых беременностях наблюдаются в несколько раз чаще, чем при обычных беременностях, например по американским данным в 5 раз (Hirst, 1940), по другим — в 2—3 раза (Gedda, 1951); эти цифры зависят от материала и способа учета абортов.
Преждевременные роды близнецов составляют значительный процент, но далеко не одинаковый для различных стран и мест (Gedda, 1951). В качестве примера могут служить данные французских исследователей (Gernez et Omez, 1938), полученные на 226 парах близнецов.
14.15% родов вовремя 35.40% » 8.5 мес. 17.70% » 8 » 10.61% » 7.5 » 5.75% » 7 » 6.63% » 6 »
Таким образом, значительное число близнецов — недоноски разной степени, что не может не сказываться на их выживании в первые месяцы после рождения. По другим данным около 30% близнецов родится вовремя (Zazzo, 1955). При близнецовых родах относительное число мертворожденных и умирающих вскоре после рождения в несколько раз превышает число гибнущих новорожденных при обычных родах, например по данным американской статистики за 1931 — 1937 гг. — более чем в 3 раза (Yerushalmy a. Sheear, 1940в; Strandskov a. Ondina, 1947). По другим данным эта разница еще больше. Так, из числа новорожденных-одиночек гибнет 3%, из числа РБ — 8%, а из числа ОБ — 21%. Если взять только родившихся вовремя или в течение 9-го месяца, то цифры гибели будут следующие: 3% для одиночек, 3% для РБ, 15% для ОБ (Zazzo, 1955). Эти цифры говорят об особой неустойчивости ОБ, что особенно бросается в глаза при сравнении с РБ, которые в этом отношении не отличаются от одиночек. Мертворожденные и гибнущие новорожденные чаще встречаются среди близнецов одного пола, чем разного; чаще гибнут оба близнеца одной пары одинакового пола, чем разного. Возраст матери на гибель близнецов, видимо, влияния не оказывает. Мертворожденные близнецы чаще оказываются мужского пола, чем женского, о чем говорят данные нескольких исследователей (Gedda, 1951; Dahlberg, 1952). Так, в Швеции, по Дальбергу, в 1938—1947 гг. мертворожденные близнецы мужского пола составляли 6.7%, а женского 5.87%, тогда как при обычных родах количество мертворожденных достигает приблизительно 2.5%. Среди ОБ мертворожденных несколько больше, чем среди РБ. Например, на основании американской статистики 30-х годов среди ОБ число пар, в которых один близнец был мертворожденным, составляло 13.2%, а среди РБ — 10.3% (Yerushalmy a. Sheear, 1940в). Эти отношения еще недостаточно изучены. После рождения особенно много близнецов погибает в первую неделю жизни, например в США 90.4% из числа всех близнецов, гибнущих в первый месяц, что составляет несколько больший процент, чем для одиночек того же возраста (Allen, 1955а). В последующие месяцы смертность близнецов резко падает. Гибель зародышей увеличивается с увеличением их числа при многоплодных беременностях; иначе говоря, относительное число умерших эмбрионов в составе троен больше, чем в составе двоен и т. д. Например, если в американском населении процент мертворожденных близнецов был в 3.5 раза больше мертворожденных одиночек, то мертворожденных среди троен было в 7.53 раза больше (Strandskov a. Ondiua, 1947). Также, по-видимому, еще больше процент младенцев из состава троен и четверен, гибнущих в первое время после рождения. Этот вопрос, однако, еще мало изучен (Miettinen, 1954). Это значит, что при многоплодных беременностях, свыше двух зародышей, легко гибнет часть зародышей или новорожденных, а иногда и все. Таким образом, случаи, когда все три близнеца выживают, являются сравнительно редкими, еще меньше шансов встретить четверню, в которой остались живы все ее члены, и исключительно мало шансов найти пятерню, сохранившуюся до взрослого возраста. Единственный пока известный случай такой пятерни — это канадские однояйцевые близнецы Дионн (рис.36).
 Мы не можем здесь входить в рассмотрение бесчисленного количества различных осложнений многоплодных беременностей и родов, также обусловливающих гибель близнецов. Краткий обзор их дает Гедда (1951). Ограничимся одним рисунком (рис.37).
Мы не можем здесь входить в рассмотрение бесчисленного количества различных осложнений многоплодных беременностей и родов, также обусловливающих гибель близнецов. Краткий обзор их дает Гедда (1951). Ограничимся одним рисунком (рис.37). В заключение надо коснуться вопроса, который естественно возникает при рассмотрении различных аномалий и трудностей пренатального развития близнецов: являются ли близнецы вообще, особенно ОБ, нормальными людьми? Несомненно, что в ряде случаев утробная жизнь близнецов оставляет на них и после рождения какой-то след. Но, во-первых, это бывает не всегда, и нередко близнецы рождаются полноценными младенцами, а во-вторых, во многих случаях, как например у недоносков-близнецов, если и наблюдаются различные аномалии после родов, то впоследствии они постепенно ликвидируются или сглаживаются, как мы это увидим из дальнейшего изложения. Поэтому на основании некоторых случаев неполноценности близнецов нельзя делать обобщающий вывод о неполноценности близнецов вообще, а также об относительной неполноценности одного из пары близнецов (Lotze, 1937; Newman, 1940в; Gedda, 1951; Verschuer, 1954, и др.).
Сказанным, однако, не исчерпывается круг вопросов, относящихся к пренатальному развитию. Существуют, оказывается, факторы, влияние которых на организм восходит ко времени, когда организм еще не существовал, когда зигота, из которой он образовался, еще не возникла. Такие факторы относятся к области «прогенеза», т. е. к доэмбриональной истории индивидуума, если можно так сказать, — это факторы, влияющие на гаметы в период их становления, до возникновения зародыша. Особенно важно это учитывать в отношении яиц, поскольку в яйце в период его созревания намечается ряд структурных моментов будущего организма, как например билатеральная симметрия и т. д. К области «прогенеза» относятся как факторы наследственные, так и ненаследственные. К первым принадлежат генотипы обоих родителей, ко вторым относятся: возраст родителей, разница в их возрасте, число детей в семье, интервалы между рождением детей, пол предшествующего ребенка, болезни родителей, алкоголизм их и т. п. (Turpin, 1955). Вся эта область только недавно стала систематически изучаться и, конечно, представляет большой интерес и для близнецовой проблемы.
В заключение надо коснуться вопроса, который естественно возникает при рассмотрении различных аномалий и трудностей пренатального развития близнецов: являются ли близнецы вообще, особенно ОБ, нормальными людьми? Несомненно, что в ряде случаев утробная жизнь близнецов оставляет на них и после рождения какой-то след. Но, во-первых, это бывает не всегда, и нередко близнецы рождаются полноценными младенцами, а во-вторых, во многих случаях, как например у недоносков-близнецов, если и наблюдаются различные аномалии после родов, то впоследствии они постепенно ликвидируются или сглаживаются, как мы это увидим из дальнейшего изложения. Поэтому на основании некоторых случаев неполноценности близнецов нельзя делать обобщающий вывод о неполноценности близнецов вообще, а также об относительной неполноценности одного из пары близнецов (Lotze, 1937; Newman, 1940в; Gedda, 1951; Verschuer, 1954, и др.).
Сказанным, однако, не исчерпывается круг вопросов, относящихся к пренатальному развитию. Существуют, оказывается, факторы, влияние которых на организм восходит ко времени, когда организм еще не существовал, когда зигота, из которой он образовался, еще не возникла. Такие факторы относятся к области «прогенеза», т. е. к доэмбриональной истории индивидуума, если можно так сказать, — это факторы, влияющие на гаметы в период их становления, до возникновения зародыша. Особенно важно это учитывать в отношении яиц, поскольку в яйце в период его созревания намечается ряд структурных моментов будущего организма, как например билатеральная симметрия и т. д. К области «прогенеза» относятся как факторы наследственные, так и ненаследственные. К первым принадлежат генотипы обоих родителей, ко вторым относятся: возраст родителей, разница в их возрасте, число детей в семье, интервалы между рождением детей, пол предшествующего ребенка, болезни родителей, алкоголизм их и т. п. (Turpin, 1955). Вся эта область только недавно стала систематически изучаться и, конечно, представляет большой интерес и для близнецовой проблемы.
Глава шестая
СОЕДИНЕННЫЕ БЛИЗНЕЦЫ
Нам уже приходилось упоминать о соединенных близнецах, представляющих собой картину как бы частично раздвоенного организма (двухголовые формы и т. п.). Такого рода явления называют еще «двойными образованиями» («Doppelbildungen», по Швальбе), «двойными уродствами», или «сросшимися близнецами». Последнее название может вызвать представление, что такие близнецы получаются путем соединения ранее независимых друг от друга зародышей, быть может, даже РБ. Согласно современным представлениям, далеко не все «соединенные близнецы» возникли путем вторичного соединения. Наоборот, в ряде случаев предполагается раздвоение ранее единого зачатка, т. е., как уже не раз упоминалось, «соединенные близнецы» являются случаями ОБ, не достигших полного разделения, не вполне обособившихся друг от друга. Поэтому в самом названии «двойные образования» не должно содержаться указания на способ их возникновения, как в названии «сросшиеся близнецы», поскольку «срастание» или «слияние», не являются единственным или универсальным способом возникновения этого рода образований. В русской литературе нет еще твердо установленного термина для всей совокупности этих явлений. Вероятно, лучше всего употреблять термин «двойные образования» или «соединенные близнецы», не предрешая при этом способа их возникновения. На этом вопросе мы подробнее остановимся в другом месте, а теперь перейдем к вопросу о классификации «двойных образований». В большинстве случаев, как мы сейчас увидим, это явления, относящиеся к области тератологии, науки об уродствах (см. Schwalbe, 1907, 1923; Hubner, 1911; Wolff, 1948; Potter, 1953, и др.). Обычно «соединенные близнецы» состоят из двух компонентов, редко — из большего числа. Сочетания и характер развития компонентов могут быть весьма разнообразны и нелегко поддаются рациональной систематизации. Все существующие классификационные таблицы, лишь в известной мере охватывая огромное многообразие двойных образований, все же очень полезны для желающих подробнее разобраться в этом сложном материале. Мы приводим таблицу Швальбе (Schwalbe, 1923), получившую широкое признание. Надо пояснить некоторые термины этой таблицы. Швальбе называет каждого из двух компонентов пары близнецов «индивидуальной частью». Латинские термины отдельных двойных уродств состоят в большинстве случаев из двух слов; первое обозначает тот участок тела, которым соединены компоненты, конец термина — pagus — (происходит от греческого слова) значит соединять, например: thoracopagus — соединенные грудью (thorax — грудь), craniopagus — соединенные черепом (cranium — череп) и т. д. Эта таблица начинается с обычных близнецов (gemini), несоединенных.
КЛАССИФИКАЦИЯ ДВОЙНЫХ ОБРАЗОВАНИИ
Свободные двойные образования. Gemini, ОБ
A. Равномерное (симметричное) развитие индивидуальных частей компонентов: gemini aequales (равные близнецы). Б. Неравномерное (асимметричное) развитие индивидуальных частей: gemini inaequales (неравные близнецы). I. Развитие асимметричного компонента в общем редуцировано. 1. Формы тела и органов еще различимы: hemiacardius. 2. Формы тела и органов вовсе не различимы: holoacardius amorphus.(Рис. 33). II. Большой участок тела верхний или нижний отсутствует полностью или почти полностью. 3. Отсутствует верхняя часть тела: holoacardius acephalus.(Рис. 32). 4. Отсутствует нижняя часть: holoacardius acormus.(Рис. 31).
Соединенные двойные образования. Duplicitates
А. Равномерное развитие индивидуальных частей: duplicita symmetros. I. Вертикальная плоскость симметрии (формы с двойной и простой симметрией). 1. Брюшное соединение. а) Надпупочное. _а) Полное надпупочное соединение: cephalothoracopagus («янус»; рис. 42). _b) Неполное надпупочное соединение: prosopthoracopagus, thoracopagus (торакопагус), sternopagus (стернопагус), xyphopagus (ксифопагус; рис. 38); (craniopagus frontalis). б) Подпупочное соединение: ileoxiphopagus. в) Над- и подпупочное соединение: ileothoracopagus; (сеphalothoracoileopagus). 2. Спинное соединение: pigopagus (пигопагус; рис. 39); (craniopagus occipitalis). II. Горизонтальная плоскость симметрии (формы с двойной и простой симметрией). 1. Черепное соединение (краниопагус): craniopagus parietalis, из него выводятся craniopagus occipitalis и cr. frontalis (рис. 40, 50). 2. Каудальное соединение: ischiopagus (исхиопагус; рис. 41). III. Срединные плоскости индивидуальных частей параллельны к плоскости симметрии или частью совпадают с ней, частью расходятся. 1. Срединные плоскости дивергируют в краниальном направлении: duplicitas anterior (рис. 43). 2. То же в каудальном направлении: duplicitas posterior (рис. 44). 3. Расхождение в средней части: duplicitas media. 4. Комбинированные формы.
Б. Неравномерное развитие индивидуальных частей: duplicitas asymmetros (autosit и parasit). Место прикрепления паразита на теле автозита. 1. В области головы: craniopagus parasiticus (рис. 45, 46, А),janus parasiticus, epignathus и родственные формы. 2. В области шеи, груди или живота: thoracopagus parasiticus (рис. 11), epigastrius. 3. В области таза: ischiopagus parasiticus, pigopagus parasiticus (рис. 46, В) и др.







Типы близнецов, представленные в этой таблице, встречаются у различных групп животных, как беспозвоночных, так и позвоночных всех классов (см. главу восьмую), включая и человека. Мы видим, что свободные ОБ входят в общую классификацию с соединенными близнецами. Этим правильно отражается общность путей и причин образования тех и других. Но на этих вопросах мы подробнее остановимся дальше, как и на несимметричных свободных компонентах (gemini inequales). Соединенные близнецы рождаются сравнительно очень редко; например, в одном чикагском родильном доме на 60 000 родов оказались всего одни роды симметрично соединенной пары и несколько асимметричных пар (Potter, 1953). Соединенные близнецы всегда одинакового пола, причем женского пола таких близнецов рождается относительно больше, чем мужского пола, приблизительно в отношении 2 : 3. Причина этого пока не выяснена (Gates, 1946). Может быть, это происходит в силу большей выносливости женского пола к трудным условиям. Теперь нам надо перейти к рассмотрению некоторых конкретных случаев соединенных близнецов, могущих служить иллюстрацией к приведенной таблице Швальбе. Некоторые аналогичные примеры на животных приводятся в главе восьмой. К самым удивительным формам двойных уродств относятся кефалоторакопаги (cephalothoracopagus), т. е. соединенные головой и грудью; их называют еще «янусы», поскольку они напоминают изображение этого римского бога (рис. 9). Эти двойные уроды известны только в эмбриональном состоянии (рис. 42). Лица их обращены в противоположные «стороны, причем в ряде случаев лицо с одной стороны более развито, чем с другой, вплоть до почти полного его исчезновения. Швальбе предлагает так представить себе строение таких «янусов»: если взрезать ножом от пупа до темени по средней линии двух субъектов, повернутых лицом друг к другу, развернуть в стороны правую и левую половину каждого и сдвинуть вплотную обоих субъектов друг к другу; тогда правая половина головы и груди одного соединится с левой другого. Лицо и грудная клетка каждого из партнеров окажутся под прямым углом к тазу и ногам. Такова структура симметричного «януса» с одинаково развитым лицом с той и другой стороны (рис. 47). Если же мысленно представить себе различные степени смещения сливающихся частей, то за счет расширения одного из лиц произойдет сжатие противоположного лица вплоть до разных степеней его редукции; это будут разные формы несимметричного «януса». Коллекция уродов этого типа имеется в Кунсткамере Академии наук СССР в Ленинграде и описана Яцутой (1913).

Как у людей, так и у других животных этот тип уродств очень редок. Описан, например, такой «янус» у свиньи, у курицы, у скорпиона и других животных (Schwalbe, 1907). Экспериментально такие уроды получены Шпеманом и его учениками на тритонах путем сращения двух полугаструл, у которых впячивание шло навстречу друг другу (см. глава восьмая «амфибии»). При меньшей степени соединения, например при соединении сравнительно небольшого участка в области груди, двойные образования могут жить после рождения, иногда даже долго, как это было с знаменитыми «Сиамскими близнецами» Чангом и Энгом, родившимися в 1811 г. в Сиаме, откуда и пошло их название, ставшее впоследствии нарицательным для разных вариантов соединенных близнецов (это о Чанге и Энге писал М. Твэн в своем рассказе «Сиамские близнецы»). Пара эта принадлежала к ксифоиагам (xiphopagus), соединенных в области мечевидного отростка грудной кости (рис. 38). Соединяющие их ткани были преимущественно наружные, образовавшие сравнительно короткий и узкий жгут, диаметром около 10 см. Вероятно, еще и в то время можно было бы разъединить этих близнецов путем несложной хирургической операции. Однако они этого не хотели, так как их уродство было источником их богатства и известности — они разъезжали по разным странам и показывались за плату. В конце жизни они поселились в США, где купили себе по имению и жили по очереди то в одном то в другом. Они женились на сестрах, и каждый имел около 10 человек детей. Жили они дружно, только в детстве будто бы поссорились из-за температуры воды в ванне, которая одному из них показалась недостаточно теплой. В характере и вкусах они несколько отличались, в частности Чанг тяготел к алкоголю. Интересно, что у них наблюдались некоторые черты «зеркальности»: у Чанга левый глаз был слабее правого, а у Энга — наоборот. Умерли они в возрасте 63 лет, в 1874 г. Чанг заболел пневмонией и умер ночью, пока его близнец спал. Энг вскоре обнаружил смерть брата, очень горевал и умер через 2 часа, хотя до того был здоров. При жизни «сиамских близнецов» их обследовали некоторые ученые (Bolton, 1830, и др.). Вскрытие их трупов родственники не разрешили, позволив изучить только соединяющую их спайку (Ahlfeld, 1880; Груздев, 1914; Gedda, 1951, и др.). Другой парой известных ксифопагов были девочки Радика и Додика, родившиеся в 1889 г. в Индии, южнее Калькутты. Их привезли в Европу для показа публике. На 13-м году жизни Додика тяжело заболела легочным туберкулезом, температура ее тела на 3° превышала температуру тела ее близнячки. Знаменитый хирург Дуайен произвел успешную операцию их разделения: обе девочки остались живы. Однако Додика вскоре все же умерла от туберкулеза, а Радика осталась жить. Спайка, соединявшая девочек, имела около 10 см в длину и около 28 см в окружности; кровеносные системы обеих сообщались, что было установлено экспериментально: Радике давали метиленовую синьку, и вскоре она обнаруживалась в моче Додики (Груздев, 1914). Удачные попытки разъединить ксифопагов были описаны еще в XVII в. С тех пор такие операции с успехом делались многократно, даже когда в смычку входил участок печени (Aird, 1954). Переходим к пигопагам. Известным примером таковых служат «богемские сестры» Роза и Йозефа Блажек(1878— 1922), родившиеся в Праге (рис. 39). Они интересны тем, что заметно отличаются друг от друга по разным свойствам. В детстве они иногда болели неодинаково, например на 13-м году Роза перенесла дифтерию, а Йозефа осталась здорова. Зато вскоре последняя, испугавшись собаки, заболела хореей (пляска Вита), продолжавшейся 13 недель. Говорить они начали на втором году. Менструации у обеих начались на 13-м году и приходили одновременно, через 4 недели. Сестры Блажек также много разъезжали по разным странам, показываясь публике за деньги. Их исследовали как при жизни, так и посмертно (Marchaut, 1881; Baudoum, 1901; Bland-Sutton, 1922; Breakstone, 1922; Perlstein, 1927; Cummins, 1936, и др.). Роза была левым компонентом, Йозефа — правым. Рост первой был 144 см, второй — 142. Обе весили 85 кг. Правая нога Розы и левая Йозефы были «внутренние» и расположены несколько вперед, так как позвоночники близнецов находились под углом 45° друг к другу. Левая нога Йозефы на 5 см была короче правой; ходили они «в ногу». Йозефа левша. Форма головы различная — у Розы грушевидная, асимметричная. Цвет радужной глаз и цвет волос одинаковые. У них было одно анальное и одно наружное половое отверстие. Общие части тела у них имели и общую чувствительность. Внутренние половые органы разные. У Йозефы матка оказалась недоразвитой, у Розы — нормальной, и в 1910 г. она родила нормального ребенка, мальчика. Лактация была у обеих. Сон и аппетит у них разные. Пульс у Розы чаще, чем у ее близнячки. Роза живее, «интеллигентнее». Она глава пары, она ведет корреспонденцию и подписывает контракты с импрессарио; Роза не раз влюблялась. Йозефа более флегматична и инертна. Близнецы нередко ссорились, а в детстве и дрались. Девочками лазали по деревьям. Общая «интеллигентность» обеих считалась «ниже нормы». Об их дактилоскопии, а также еще двух других пар пигопагов — см. главу десятую «кожа». Из живших в прошлом столетии пигопагов надо отметить пару Милли и Кристина, родившихся в 1851 г. у невольницы-негритянки в США и воспитанных плантатором Смитом. Эти близнецы искусно танцевали и пели, притом будто бы разными голосами: одна сопрано, а другая контральто. Они получили прозвище «двухголовый соловей» (Быховский, 1927, и др.).

Изучались и другие взрослые пигопаги, жившие в наше время: Мери и Маргарет Джиб, Дэзи и Виолет Хилтон, Луцио и Симплицио Годена. Последняя пара была соединена подвижно (рис. 48). Все они имели внутрипарные различия в том же роде, как и «богемские сестры», но менее резко выраженные. В общем же разница внутри этих пар больше, чем у несоединенных ОБ. Ньюмен объясняет это тем, что соединенные близнецы возникли на более поздней стадии развития, чем обычные ОБ, когда уже наметились правая и левая стороны зародыша; таким образом, соединенные близнецы возникли один из правой, другой из левой половины с соответствующими различиями и элементами «зеркальности» (Newman, 1931, 1940b, и др.). Заметные внутрипарные различия, аналогичные найденным у людей, описаны и у соединенных близнецов домашних животных (Keller u. Niedoba, 1937).Подробнее мы к этому вопросу еще вернемся в главе восьмой.

Переходим к краниопагам, близнецам, соединенным головой. Редчайшие случаи этих двойных уродств описаны еще начиная с XVI в. Так, близ Вормса в Германии в 1495 г. родились две девочки «в общем благообразные, однако от темени до лба сросшиеся между собой и смотрящие друг на друга», как писал о них некий Мюнстер в 1544 г. (рис. 49). В возрасте 10 лет одна из них умерла. Живую отделили от мертвой, но и вторая вскоре умерла. К. Бэр (1845б) приводит цитату из Мюнстера, описание с рисунком другой пары краниопагов, соединенных теменем, «двойного ребенка» из Брюгге, и описание еще третьей пары, сделанное Анелем в 1716г. (см.: Канаев, 1951). Сам Бэр изучил анатомию пары симметричных краниопагов из коллекции Академии наук (рис. 50). Черепа их были соединены в правой лобной области и симметрично деформированы. Большое отверстие соединяло полости обоих черепов, правые доли больших полушарий сливались, передняя часть головного мозга была слабо развита. Другую пару детей краниопагов Бэр наблюдал живыми и описал различия их внешности и поведения (1856). Ткани их мозга в месте соединения сливались (Наранович, 1856). Недавно обнаружена пара краниопагов — младенцев Смит из Тасмании, родившихся в 1950 г. (Gedda, 1951). Другая аналогичная пара привлекла внимание всей Европы в связи с вопросом об оперативном разделении таких близнецов (Durr, 1952).
К группе, родственной по симметрии к краниопагам, относятся каудально соединенные близнецы — исхиопаги (ischiopagus) (рис. 41). Такие уродцы редко выживают после рождения. Стоять они не могут, так как ноги у них не развиты. Одна такая пара, названная Даша-Маша и живущая в одной из клиник в Москве, достигла к 1955 г. уже 6 лет. Она находится под постоянным наблюдением изучающих ее специалистов. Переходим к типу duplicitas parallelа, где срединные плоскости компонентов до известной степени параллельны общей плоскости симметрии. Среди форм этого типа относительно чаще встречается форма duplicitas anterior, т. е. раздвоенная спереди. Редко встречаются формы с очень незначительным раздвоением головы (рис. 14, А и рис. 51), чаще имеется две головы, иногда два торса и т. д., тогда как нижняя часть тела не раздвоена — таз один и одна пара ног (рис. 52). Существует сообщение,что при дворе шотландского короля Якова IV (1488— 1513) жил такой урод мужского пола, имевший верхнюю часть тела двойную и две пары рук, но одну пару ног. Он достиг 28 лет. Обе головы его имели заметно разную психику, и компоненты часто ссорились. Эти близнецы получили хорошее по тому времени воспитание, знали несколько языков, занимались музыкой и т. д. В 1829 г. в Париже была известна аналогичная пара младенцев - Рита и Кристина, анатомия которых изучалась (Груздев, 1914; Gedda, 1951, и др.). Рига, правая, была более слабой, имела зеркальное расположение внутренностей — situs inversus viscerum (см. главу седьмую) и аномалии кровяносной системы. В конце прошлого века жили такого же типа соединенные близнецы Точчи, родившиеся в 1877 г. в Италии и достигшие юношеского возраста (рис. 43). Выше шестого ребра они были как бы два человека, а ниже — один. Правый был лучше развит и умственно стоял выше левого. Общими у них были лишь прямая кишка, анус и пенис. Одной из двух ног двигал один близнец, второй ногой — другой (Harris, 1892; Gedda, 1956, и др.).


В Москве в 1930-е годы была подробно изучена подобная пара женского пола Ира-Галя (рис. 53; Анохин, 1939; Black, 1940). Эти дети жили немного дольше года. Их развитие и поведение наблюдали с самого рождения, а после смерти изучили их анотомию. С них снят содержательный кинофильм. Материалы исследований полностью еще не опубликованы. У этой пары позвоночник был двойной до таза, но самый таз был один и одна пара ног. Центральная нервная система у каждого компонента была почти полностью обособлена. Реакции и поведение каждой близнячки были в значительной мере самостоятельными, например: боль от укола каждая чувствовала отдельно от другой, одна спала в то время, как другая играла, и т. д. Нечто подобное наблюдал и Бэр на описанной им паре краниопагов. Несмотря на то, что кровеносные системы Иры-Гали сообщались и состав крови их в общем был одинаков, сон у них, как отмечалось, не всегда совпадал, что говорит в пользу корковой теории сна И. П. Павлова. Работа сердца, желудка и т. д. шла у каждого компонента своим ритмом (Алексеева, 1941).

Нам остается еще кратко рассмотреть некоторых представителей второго раздела (Б) таблицы Швальбе — несимметричные соединенные образования (duplicitas asymmetros). Основная особенность этой группы заключалется в том, что один из компонентов достигает более или менее полного развития (автозит), тогда как другой в той или иной мере отстает и может жить только за счет своего близнеца (паразит). Один из самых удивительных случаев этого рода был обнаружен в Индии. Это был в общем нормальный ребенок, родившийся 1783 г., на голове которого находилась живая паразитическая голова его недоразвившегося рассосавшегося в материнской утробе близнеца (рис. 45). Рот паразитической головы мог производить сосательные движения и выделять слюну: нос выделял слизь, зрачек сохранившегося глаза (другой пострадал во время родов) мог сокращаться на свету. Во время сна ребенка глаза паразитной головы не были вполне закрыты. Когда ребенок плакал, голова-паразит тоже морщилась и как бы плакала. Когда ребенок сосал, то и паразитическая голова будто бы выражала удовольствие и слюна у нее выделялась обильнее, чем обычно. Когда щипали паразитическую голову, ребенок, видимо, этого не чувствовал или меньше чувствовал, чем когда щипали его собственную голову. Уши, язык и нижняя челюсть паразита были недоразвиты, но зубы появились. Ребенок с паразитической головой был зарисован в возрасте около 2 лет; вскоре он погиб от укуса змеи. Череп его был доставлен в Англию анатому Хантеру (Home, 1790). В редких случаях автозит, имеющий крупного паразита, достигает зрелого возраста. Примером может служить генуезец Коллоредо (рис. 11), родившийся около 1620 г. и описанный в книге Лицетуса (Licetus, 1665). Паразит висел на туловище автозита головой вниз, имел две недоразвитые руки, каждая с тремя пальцами, и недоразвитую левую ногу. Из открытого рта паразита текла слюна, глаза были закрыты, он иногда слегка шевелил губами. Другой случай описан в XIX в. Это индус Лалоо, на груди которого находился крупный паразит, лишенный головы, но имеющий четыре конечности. Заднего прохода паразит не имел, но имел пенис, из которого временами выделялась моча. Лалоо прожил свыше 30 лет и был якобы женат (Груздев, 1914, и др.). Редкие случаи других асимметричных уродов описаны в тератологической литературе (Швальбе и др.) и представляют специальный интерес, почему мы здесь на них останавливаться не будем. В ряде случаев близнец-паразит деградирует до бесформенной массы тканей, иногда содержащей зуб или участок недоразвитого органа, и образуя род опухоли на теле автозита (рис. 46). Такие опухоли удаляются иногда хирургическим путем, как и более развитые паразиты. Успех такой операции прежде всего зависит от местоположения паразита и степени его слияния с какими-нибудь важными органами автозита. Тройни, соединенные вместе, очень большая редкость (рис. 54). На изображенном уроде видно, что сначала произошло раздвоение переднего конца эмбриона, а затем повторное раздвоение левой (на рисунке) головы. Вскрытие показало, что урод имел 3 гортани, 3 трахеи и 3 пищевода, но 2 пары легких, 2 сердца, 2 позвоночника, из которых один раздвоился вторично только в самом переднем конце. Эта редкая тройня аналогична тройням рыб, изображенным на рис. 55. Некоторые другие тройни изображены в специальных работах (Ahlfeld, 1880; Schwalbe, 1907, и др.).


У животных подобные уродства также представляют большую редкость. Например, описан тройной ягненок, родившийся мертвым. Он состоял из основного, более близкого к норме компонента и двух придаточных, только частично развитых. Голова и шея были нормальные и одиночные. Грудь двойная, 2 пары передних конечностей. Позвоночников было 2 и 2 хвоста, но неравные по размерам. Задних конечностей было 3 пары. Третья пара ног имела рудимент таза и являлась типичным паразитом (Lesbre et Tagant, 1927). Соединенные четверни, разумеется, встречаются еще реже, чем тройни. По-видимому, компоненты в этих случаях не развиваются нормально, и большинство их оказывается паразитами. В качестве примера можно привести исключительный случай (рис. 56), описанный в прошлом веке и приведенный Швальбе (Schwalbe, 1907). У первого компонента в области рта прирос бесформенный второй компонент, а к нему на раздвоенной пуповине — остатки двух других компонентов, сведенных каждый к недоразвитому тазу с парой ног. Все четыре компонента женского пола, очевидно однояйцевого происхождения. Урод этот погиб в качестве выкидыша на пятом месяце беременности.

Вопрос о способах возникновения соединенных близнецов до сих пор нельзя еще считать вполне выясненным, несмотря на ряд гипотез в этой области, которых выше мы уже не раз касались. Попытка унифицированного объяснения всевозможных форм двойных уродств Грэпером (Graper, 1931) путем сведения их к разнообразным столкновениям «потоков» тканей в период ранней гаструляции, по-видимому, ошибочна. Вероятно, разные типы соединенных близнецов возникают на различных стадиях развития и разными путями (Politzer, 1955). Прежде всего надо подчеркнуть, что соединенные близнецы являются, очевидно, всегда однояйцевыми. В пользу этого допущения говорит целый ряд фактов. Оба компонента всегда одного пола. У них всегда одинаковы группа крови и факторы крови. Амнион у них один общий. Часто они соединены гомологичными местами груди, черепа и т. д., и компоненты симметрично расположены по отношению друг к другу. Это последнее обстоятельство лежит в основе классификации двойных уродств. Нет ни одного достоверного факта, говорящего в пользу допущения срастания двух эмбрионов, возникших из разных яиц, и образования таким путем двойных уродств (Е. Wolff, 1948, и др.). Экспериментально можно, как известно, достигнуть слияния двух зигот или двух зародышей с образованием гармоничного зародыша удвоенного размера или «сросшихся близнецов», например у морских ежей (Driesch, 1903, и др.) или амфибий (Боги, 1897; Mangold u. Seidel, 1927; Е. Wolff, 1948, и др.). Но это возможно только при удалении оболочек и при других экспериментальных вмешательствах в процессе развития, т. е. таких изменениях его, которые в естественных условиях произойти не могут. В дальнейшем мы на конкретном материале познакомимся с экспериментальным получением соединенных близнецов у разнятых форм животных, начиная от низших, и объяснениями отдельных случаев такого рода. Здесь же мы лишь кратко остановимся на некоторых общих представлениях об этих явлениях, дополнительно к вышесказанному о причинах возникновения ОБ. Согласно новейшим представлениям (Politzer, 1955), соединенные близнецы могут возникать по крайней мере тремя путями: 1) во время ранней гаструляции происходит раздвоение переднего конца зародыша; 2) происходит «слияние» тканей в период гаструляции двух смежных зародышей (ОБ) еще до дифференцировки органов; 3) происходит «срастание» двух зародышей (ОБ) уже после известной дифференцировки органов, после гаструляции. Рассмотрим эти три способа образования двойных уродств. Переднее раздвоение, с которым мы уже не раз встречались, как известно, может быть также получено экспериментально, например перетяжкой на яйцах тритонов. В этих опытах те части тела, которые оказались раздвоенными, получают приблизительно около половины нормально полагающегося материала и оказываются в связи с этим меньшего размера против нормы, а кроме того, иногда еще с дефектами развития (циклопия и т. п.). В случаях образования переднего раздвоения в естественных условиях, т. е. без экспериментального вмешательства, удвоенные передние части тела имеют нормальную величину («закон» Тура, 1903), чем и отличаются от переднего раздвоения, полученного в опыте. Политцер считает, что этим можно объяснить возникновение раздвоения в естественных условиях: вероятно, в силу каких-то физиологических причин (пока не выясненных) подлежащего инвагинации при гаструляции материала оказывается больше, чем полагается, и раздвоение его происходит в порядке регуляции. Хотя закон Тура наблюдается у разных форм животных (амфибий, птиц, человека), однако объяснение его как результат «регуляции» вызывает сомнение и требует более убедительного обоснования, чем у Политцера. Остается неясным также самый «механизм» такого раздвоения при инвагинации. Политцер допускает разные варианты его: или образование двух смежных бластопоров с последующим слиянием тканей внутри гаструлы, как это описано на одном яйце ящерицы (Kopsch, 1897), или раздвоение сначала нормальной инвагинации по пока не выясненной причине. Заднее раздвоение, обычно встречающееся гораздо реже переднего, вряд ли может быть объяснено подобным образом; Политцер предполагает, что этот род раздвоения можно объяснить «срастанием». Второй способ образования соединенных близнецов — это «слияние» двух смежных зародышей, возникших в одном яйце. В зависимости от их расположения по отношению друг к другу и степени их близости слияние их может происходить в различных участках тела зародышей и в разной степени. Так, например, при росте двух зародышей, расположенных радиально и под значительным углом (до 180°), они могут наткнуться друг на друга головными концами, причем ткани их могут при этом слиться. Примером могут служить нижеприведенные зародыши кур (рис. 101, 103).


При более или менее параллельном расположении осей тела зародышей, что бывает при сравнительно близком их нахождении друг от друга в яйце, зародыши в процессе развития и роста приближаются друг к другу в связи с перемещением тканей внутрь гаструлы. При этом зародыши сближаются настолько, что ткани их в той или иной мере сливаются, образуя впоследствии общие органы, так что невозможно определить, какая часть такого общего органа произошла из тканей того или другого компонента.Таково происхождение «янусов», таракопагов, исхиопагов и тому подобных двойных уродств. Обращает на себя внимание симметричность таких уродов, порой удивительно ясно выраженная. При соединении обоих зародышей, очевидно, происходит слияние подобных зачатков органов между собой: сомитов с сомитами, сердца с сердцем, печени с печенью и т. д., т. е. проявляется тенденция слияния «подобного с подобным», отмеченная еще Э. Жоффруа Сент-Илером. Физиологический механизм этих явлений пока не изучен. По-видимому, симметрия соединенных близнецов этой группы находится в зависимости от этой тенденции — сродства «подобного к подобному». Близким по существу с изложенным способом образованием соединенных близнецов является третий способ — «срастание»: это соединение двух ОБ на сравнительно более поздней стадии развития, когда органы зародышей уже более развиты. Примером могут служить некоторые краниопаги, где компоненты соединены негомологичными местами и потому расположены не вполне симметрично друг к другу (ср. рис. 40). В месте соединения ткани обоих близнецов могут тоже до известной степени смешиваться и образовывать общий обоим участок тела, но в общем меньший по размерам и более периферийный, чем у некоторых соединенных близнецов второй группы. Возможно, что и некоторые асимметрично соединенные пигопаги возникают путем срастания. Трудно, конечно, провести отчетливую грань между явлениями слияния и срастания зародышей. Политцер вовсе не выделяет продольное расщепление зародыша как способ образования соединенных близнецов такого рода, как заднее раздвоение и другие. Вообще весь вопрос образования соединенных близнецов, как и ОБ вообще, еще далеко не достаточно изучен и потребует дальнейшего собирания материала и исследования.
Изучение соединенных близнецов затрагивает ряд интересных вопросов эмбриологии, на которых мы здесь останавливаться не можем. Но одному вопросу, выходящему за рамки эмбриологии и имеющему большой теоретический интерес, мы посвящаем особую главу — это симметрия, асимметрия и зеркальность близнецов как соединенных, так и несоединенных, «свободных».
Глава седьмая
ВОПРОСЫ СИММЕТРИИ, АСИММЕТРИИ И ЗЕРКАЛЬНОСТИ У БЛИЗНЕЦОВ
Как известно, большинство классов животных, близнецы которых изучались, имеет билатеральное, двусторонне-симметричное строение тела (черви, насекомые и другие членистоногие, все хордовые и отчасти иглокожие). Иначе говоря, у всех этих животных можно различить правую и левую стороны тела, в известной мере симметричные друг другу. Так, например, у позвоночных животных правый глаз симметричен левому, также уши, конечности и т. д.; из внутренних органов — почки, яичники, семенники, легкие и т. д. Тело такого животного можно мысленно рассечь плоскостью симметрии на правую и левую половину, и каждая из них будет как бы зеркальным отражением другой. Каждой точке тела, расположенной по одну сторону плоскости симметрии,соответствует такая же точка на другой стороне этой плоскости, и обе симметричные точки находятся на равном расстоянии от плоскости симметрии. Однако при ближайшем рассмотрении животных оказывается, что симметрия их в той или иной форме и степени оказывается нарушенной, иногда очень резко. Это явления асимметрии. Так, у человека сердце, желудок, печень и другие органы расположены асимметрично: сердце влево от плоскости симметрии, печень вправо и т. д. Описаны различия правой и левой стороны головного мозга, обоих легких, сосудистой системы, кожных борозд пальцев и ладоней, пигментации волос и многие другие различия но размерам, форме и функции частей правой и левой стороны. При этом можно различать асимметрии нормальные, как расположение сердца слева, а желудка справа, и ненормальные, когда обычно симметричные части оказываются асимметричными, например одна половина носа или пениса больше, чем другая — явления гемигипертрофии (Gesell, 1921, 1927; Verschuer, 1932, и др.), или только одна половина тела пигментирована (Злотников, 1945). Но и в структурах, на первый взгляд кажущихся нормальными, симметричными, асимметрия может быть легко обнаружена. Так, например, сравнение лица пары ОБ позволяет видеть и здесь асимметрию, особенно, если по-разному комбинировать правую и левую половину лица того же субъекта или обоих близнецов (рис. 57; Freerksen, 1938, и др.). Морфологофизиологическая асимметрия животного мира бесконечно разнообразна. Биологическое значение этих явлений и их происхождение еще мало выяснены (Ludwig, 1932, 1949; Child, 1941, и др.).

Эта проблема асимметрии, или левизны-правизны, как ее еще называют, не ограничивается животным миром. Асимметрию можно обнаружить в морфологии растений и микроорганизмов (Алпатов, 1957), в строении протоплазмы всего органического мира (Гаузе, 1940), в мире кристаллов, в строении молекул и атомов (Вернадский, 1940). Асимметрия обнаруживается и за пределами нашей планеты и встает в качестве астрономической проблемы при изучении спиральных туманностей и других явлений в мировом пространстве. Мы, разумеется, не можем пытаться здесь разобраться в целом в этой большой и очень еще мало понятной проблеме. Мы можем лишь остановиться на тех фактах, которые непосредственно связаны с изучением близнецов. Как известно, при сравнении пары близнецов можно различать три рода структурного сходства: первый — это билатеральное сходство, т. е. сходство правой и левой стороны у обоих; второй — гомолатеральное сходство, когда одна и та же сторона, например правая, у близнецов более похожа, чем правая и левая стороны каждого из них; наконец, третий род — гетеролатеральное сходство, или зеркальное, когда правая сторона первого близнеца больше похожа на левую второго, чем на левую первого, и т. д.; в таких случаях один близнец является как бы зеркальным отражением другого. Описан ряд асимметричных признаков, как морфологических, так и функциональных, на которых проявляется такая зеркальность. Мы рассмотрим некоторые из них. 1) Завиток волос на темени. Он может идти у одного близнеца по часовой стрелке, а у другого — наоборот. 2) Узоры кожных бороздок на дистальных фалангах пальцев, на ладонях и ступнях (см. главу десятую «кожа»). Так, у одного из ОБ на правом указательном пальце имеется дактилоскопический рисунок, называемый радиальной петлей, состоящий из 5 бороздок, а у его партнера такой же рисунок на левом указательном пальце. На другой паре указательных пальцев той же пары близнецов другой рисунок — ульнарные петли из 13 бороздок (Cockayne, 1940) и т. д. Так же зеркально оказываются иногда расположенными рисунки ладоней и подошв близнецов (Wilder, 1904; Newman, 1930, и др.). 3) Расположение зубов, их аномалии и их прорезание. Например, у пары младенцев ОБ прорезание молочных зубов по местоположению и последовательности шло зеркально (Steinmann, 1943; Gedda, 1951, и др.). 4) Глаза могут быть зеркально преобладающими: у одного близнеца лучше видит левый глаз, у другого — правый. Такая зеркальность глаз может иногда совпадать с преобладанием руки той же стороны, а иногда и ноги. Описаны случаи, когда один из близнецов косит правым глазом, а другой — левым (Steinmann, 1943, и др.). Встречается также зеркальность особенностей строения лица, ушной раковины и других мелких морфологических особенностей. 5) Леворукость привлекала внимание многих ученых (ср. Gedda, 1951), которые стремились выяснить преимущественно следующие вопросы:
а) как часто встречаются левши среди населения, б) как часто встречаются левши среди близнецов, в) существует ли связь между леворукостью и другими асимметричными признаками, которые у близнецов могут проявляться зеркально, как например завиток на темени, идущий то по ходу часовой стрелки, то против него, или некоторые кожные узоры кончиков пальцев и т. д., и, наконец, г) наследственна ли леворукость.Выяснилось, что среди разных групп населения Европы, по-видимому, в среднем несколько чаще встречаются левши мужчины, чем женщины, например 4—5% мужчин и только 2—2.5% женщин по Людвигу (Ludwig, 1932). Но родится левшей, конечно, больше, так как значительный процент их путем воспитания становится правшами. Людвиг считает, что рождается около 25% левшей. Другие авторы указывают иные цифры для числа левшей среди населения, например Сименс (Siemens, 1924) приводит данные некоторых авторов, из которых явствует, что в разных группах населения, в том числе и у школьников, число левшей колеблется от 1 до 18%. У японских школьников (Komai a. Fukuoka, 1934b) леворукость обнаруживалась в разной степени в зависимости от приема испытания ее и оказалась максимальной: у мальчиков от 5 до 8.5%, у девочек от 4 до 6%. Леворукость убывает с возрастом. Райф (Rife, 1940) среди студентов-зоологов Университета Охайо (США) и их семей нашел 7.45% левшей, а по данным других исследователей среди разных групп населения — от 6,5 до 7.6%. Кроме небольших реальных различий из-за возраста и других причин среди отдельных групп населения разных стран, материал исследователей дает неодинаковые результаты и порой трудно сравним, потому что леворукость оценивается путем разных приемов, берутся разные по возрасту группы и т. д.; так, например, Райф (Rife, 1943) пользуется 10 тестами (бросать, резать ножом, шить, писать и т. п.), Комаи и Фукуока (Komai a. Fukuoka, 1934b) — семью, а другие еще меньшим числом. Что касается состава испытуемых, то Комаи, например, брал детей школьного возраста, а возраст материала других исследователей порой и вовсе не указан, например у Райфа. Наконец, само понятие «левша» довольно неопределенно, так как проявление леворукости имеет ряд степеней и нюансов и связи с различием индивидуальных навыков и т. д., что затрудняет оценку этого «признака». Но независимо от этих расхождений данные нескольких авторов показывают, что в семьях, где оба родителя левши, около половины детей оказывается тоже левшами; если же один родитель левша, то около 17% детей левши, а если оба родителя правши то левшей среди детей оказывается лишь около 6% (Rife, 1943, и др.). Это как будто говорит за то, что леворукость наследственна. Однако леворукость не наследуется как простой признак, и вопрос о характере наследования леворукости настолько неясен, что некоторые исследователи, например Сименс (Siemens, 1924) и Фершюр (Verschuer, 1932), прямо отрицают наследственность леворукости. Казалось бы, раз родословный метод не решает вопроса о передаче потомству леворукости, то не поможет ли исследование близнецов в этом деле, почему близнецов в этой связи многократно изучали. Оказалось, что леворукость чаще встречается среди близнецов, чем в массе населения, причем как среди ОБ, так и РБ, главным образом за счет того, что один, а не оба близнеца, левша. Так, например, Райф (Rife, 1940) приводит следующие данные (табл. 7), где П-П значит пара правшей, П-Л — правша-левша, а Л-Л — оба левши. Братья-сестры взяты попарно случайно.
 Аналогичные данные можно найти и у других исследователей (Verschuer, 1932; Komai a. Fukuoka, 1934b; Dahlberg, 1948, и др.), причем обращалось внимание на то обстоятельство, что левшей среди ОБ больше, чем среди РБ, и что процент пар, где один партнер левша приблизительно один и тот же как для ОБ, так и для РБ. Причина этого, а также большого числа левшей среди близнецов по сравнению с массой населения — вопросы невыясненные. Высказано несколько гипотез, нуждающихся в доказательствах. Например, Райф (Rife, 1940) предполагает, что могут быть субъекты в смысле леворукости промежуточной наследственности, т. е. гетерозиготы, и тогда при раздвоении яйца с такого рода наследственностью утробные влияния могут служить стимулом для превращения одного из ОБ в левшу, а другого в правшу. А для РБ к факторам утробной среды присоединяется еще различие в наследственности.
Нередко леворукость сочетается с «левоногостью» и лучшим видением левым глазом. Эти связи пока еще недостаточно изучены. Иногда леворукость сочетается также с направлением завитка волос на темени против часовой стрелки, тогда как этот завиток часто идет по ходу часовой стрелки; а кроме того, с некоторыми особенностями кожных узоров на копчиках пальцев и на ладонях и другими морфологическими и функциональными проявлениями «зеркальности» (Turpin et al., 1938, 1943, и др.). Это давало повод некоторым авторам (Newman, 1923—1940, и др.), как уже говорилось выше, усматривать и такого рода «зеркальности» проявление первоначальной дифференцированности на правую и левую половину того амбриона, из которого получились такие «зеркальные» близнецы; этим, казалось, и объясняется их происхождение. Но подобное объяснение не применимо к РБ. Далее, не удалось доказать, что такие признаки, как завиток волос против часовой стрелки, встречается чаще среди близнецов, чем среди массы неблизнецов (Komai a. Fukuoka, 1934а). То же надо сказать и относительно кожных узоров (Rife a. Cummins, 1943) и предельной формы проявления зеркальности — situs inversus viscerum (Torgersen, 1950).
Установлена любопытная связь леворукости с пониженными числовыми показателями кожных узоров как у одиночных субъектов, так и у близнецов-левшей, когда оба партнера левши. Достоверно это установлено только для женщин. Этой разницы нет, когда один из партнеров ОБ левша, а другой правша, что толкуется в том смысле, что леворукость одного из ОБ вызвана утробными условиями (Rife, 1943).
Ставился вопрос также о связи леворукости с расстройством речи, эпилепсией и другими аномалиями (Siemens, 1924). Такого рода связи, если они действительно существуют, пока еще мало исследованы и носят, возможно, чисто функциональный характер. Наконец, как не раз указывалось в литературе, внутрипарные различия психики ОБ могут зависеть от того, что один из них левша (Hutter, 1953). Но это вопрос еще мало изученный.
6) Зеркальное (обратное) расположение внутренностей — situs inversus viscerum (в дальнейшем s. i.) — очень редкое явление, которое выражается в том, что асимметричные внутренние органы полости тела оказываются расположенными зеркально в отношении нормы: сердце справа, печень слева и т. д. Ньюмен считает s. i. крайней степенью выражения зеркальности (рис.58) .
Аналогичные данные можно найти и у других исследователей (Verschuer, 1932; Komai a. Fukuoka, 1934b; Dahlberg, 1948, и др.), причем обращалось внимание на то обстоятельство, что левшей среди ОБ больше, чем среди РБ, и что процент пар, где один партнер левша приблизительно один и тот же как для ОБ, так и для РБ. Причина этого, а также большого числа левшей среди близнецов по сравнению с массой населения — вопросы невыясненные. Высказано несколько гипотез, нуждающихся в доказательствах. Например, Райф (Rife, 1940) предполагает, что могут быть субъекты в смысле леворукости промежуточной наследственности, т. е. гетерозиготы, и тогда при раздвоении яйца с такого рода наследственностью утробные влияния могут служить стимулом для превращения одного из ОБ в левшу, а другого в правшу. А для РБ к факторам утробной среды присоединяется еще различие в наследственности.
Нередко леворукость сочетается с «левоногостью» и лучшим видением левым глазом. Эти связи пока еще недостаточно изучены. Иногда леворукость сочетается также с направлением завитка волос на темени против часовой стрелки, тогда как этот завиток часто идет по ходу часовой стрелки; а кроме того, с некоторыми особенностями кожных узоров на копчиках пальцев и на ладонях и другими морфологическими и функциональными проявлениями «зеркальности» (Turpin et al., 1938, 1943, и др.). Это давало повод некоторым авторам (Newman, 1923—1940, и др.), как уже говорилось выше, усматривать и такого рода «зеркальности» проявление первоначальной дифференцированности на правую и левую половину того амбриона, из которого получились такие «зеркальные» близнецы; этим, казалось, и объясняется их происхождение. Но подобное объяснение не применимо к РБ. Далее, не удалось доказать, что такие признаки, как завиток волос против часовой стрелки, встречается чаще среди близнецов, чем среди массы неблизнецов (Komai a. Fukuoka, 1934а). То же надо сказать и относительно кожных узоров (Rife a. Cummins, 1943) и предельной формы проявления зеркальности — situs inversus viscerum (Torgersen, 1950).
Установлена любопытная связь леворукости с пониженными числовыми показателями кожных узоров как у одиночных субъектов, так и у близнецов-левшей, когда оба партнера левши. Достоверно это установлено только для женщин. Этой разницы нет, когда один из партнеров ОБ левша, а другой правша, что толкуется в том смысле, что леворукость одного из ОБ вызвана утробными условиями (Rife, 1943).
Ставился вопрос также о связи леворукости с расстройством речи, эпилепсией и другими аномалиями (Siemens, 1924). Такого рода связи, если они действительно существуют, пока еще мало исследованы и носят, возможно, чисто функциональный характер. Наконец, как не раз указывалось в литературе, внутрипарные различия психики ОБ могут зависеть от того, что один из них левша (Hutter, 1953). Но это вопрос еще мало изученный.
6) Зеркальное (обратное) расположение внутренностей — situs inversus viscerum (в дальнейшем s. i.) — очень редкое явление, которое выражается в том, что асимметричные внутренние органы полости тела оказываются расположенными зеркально в отношении нормы: сердце справа, печень слева и т. д. Ньюмен считает s. i. крайней степенью выражения зеркальности (рис.58) .
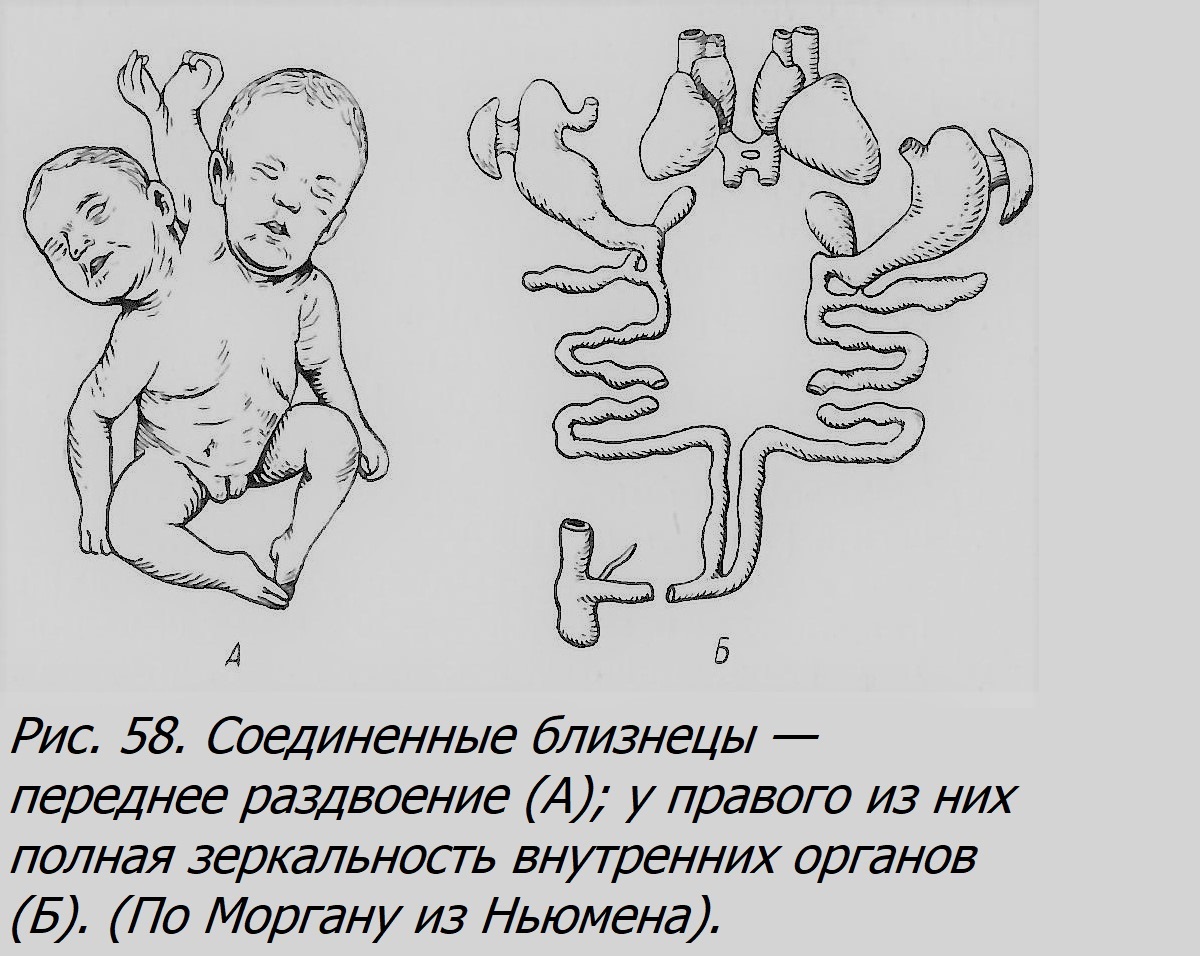 Это зеркальное расположение может быть в одних случаях полным, т. е. охватывать все внутренние органы, в других случаях частичным — касаться только сердца (декстрокардия; Stadler, 1955, и др.) и только органов грудной полости, или наоборот — только органов брюшной полости. Это неполный s. i. Такое удивительное явление уже давно обращало на себя внимание анатомов и описывалось еще в XVII в. (Gates, 1946). На основании различных исследований (Newman, 1940; Cleveland, 1945; Torgersen, 1950; Lowe a. McKeown, 1954, и др.) можно считать, что s. i. встречается приблизительно в количестве одного случая на 7 — 12 тысяч человек, т. е. очень редко. Есть указания на наследственность s. i. на основании следующих данных:
Это зеркальное расположение может быть в одних случаях полным, т. е. охватывать все внутренние органы, в других случаях частичным — касаться только сердца (декстрокардия; Stadler, 1955, и др.) и только органов грудной полости, или наоборот — только органов брюшной полости. Это неполный s. i. Такое удивительное явление уже давно обращало на себя внимание анатомов и описывалось еще в XVII в. (Gates, 1946). На основании различных исследований (Newman, 1940; Cleveland, 1945; Torgersen, 1950; Lowe a. McKeown, 1954, и др.) можно считать, что s. i. встречается приблизительно в количестве одного случая на 7 — 12 тысяч человек, т. е. очень редко. Есть указания на наследственность s. i. на основании следующих данных:
а) увеличения числа случаев его при родственных браках, б) также среди лиц, имеющих братьев-сестер с этой аномалией, в) благодаря сочетанию s. i. с некоторыми другими аномалиями (сердца, легких и др.) наследственного характера (Torgersen, 1950: Lowe a. McKeown, 1954).Среди животных, как диких, так и домашних, s. i. очень малоизучен и, по-видимому, тоже редок. Иную картину частоты представляет s. i. среди близнецов. Не останавливаясь на беспозвоночных (морских звездах,— Newman, 1923, 1940b, и др.), мы сразу обратимся к s. i. у близнецов позвоночных. В этом отношении очень показательны работы на рыбах (рис.84),
 особенно сделанные сравнительно недавно — Комаи (Komai, 1938) и Линна (Lynn, 1946), работавших на лососевых разных видов. Комаи изучил s. i. у 502 пар «свободных» ОБ, связанных только желточным мешком, и у 287 пар двойных уродств, в большинстве с передним раздвоением. У «свободных» близнецов он нашел 17.5% особен с s. i. при 6.5% у одиночек. У деформированных особей свободных близнецов s. i. наблюдался в 3 раза чаще, чем у нормальных, т. е. обнаружена корреляция между деформацией и s. i. У соединенных близнецов правый компонент был чаще деформирован и недоразвит, чем левый, и s. i. правых близнецов оказался у 66 пар, тогда как s. i. левых близнецов был только у 31 пары, т. е. в 2 раза реже.
С еще большим материалом работал Линн. Он изучил 2100 близнецов (свободных и соединенных) и 1110 одиночных особей, обращая внимание на положение и форму желудка, печени и плавательного пузыря. Одиночные особи в материале Линна имели s. i. в 4.7%. Больший процент (29.5) с s. i. был среди особей с искривлением тела в одну сторону. Интересно, что s. i. чаще встречается у особей с таким искривлением тела влево (44.7%), чем вправо (14.3%).
Среди близнецов s. i. встречался значительно чаще, чем среди одиночных особей — 37.2%. Соединенные близнецы имели s. i. чаще (41.1%), чем свободные (30.8%). В группе асимметричных соединенных близнецов, где один из партнеров оказывается недоразвитым или дефективным, паразитом, s. i. чаще встречается именно у такого близнеца, а не у более развитого (автозита). Интересно, что и по данным Линна правый близнец у соединенных имеет s. i. вдвое чаще, чем левый (22.3 % к 11.5%). Иное отношение, однако, найдено у швейцарской форели (Bovet, 1931), правда, на сравнительно небольшом материале (42 пары): у правых компонентов s. i. обнаружен в меньшем числе, чем у левых (на 9 правых компонентов 15 левых). Единичные случаи s. i., у левого близнеца описаны также у амфибий (Schwind, 1942).
Интересно, что встречаются такие случаи, когда оба свободных близнеца имеют s. i. Среди соединенных близнецов это явление наблюдается чаще (8% по Бове и 7.3% по Линну). Место соединения также влияет на частоту s. i. Чаще всего он встречается у близнецов, соединенных на уровне спинного плавника, т. е. приблизительно посередине тела.
Линн (Lynn a. Peadon, 1949) изучал и троен тех же рыб (рис.55).
Оказалось, что s. i. у одного и более компонентов тройни встречается еще чаще, чем у двоен, а именно в количестве 70.9%.
Б общем, очевидно, существует тесная зависимость между процессом образования близнецов, возникновением дефектов развития и зеркальностью внутренних органов (s. i.). Физиологический механизм этого явления еще мало выяснен. К существующим теоретическим интерпретациям зеркальности близнецов мы еще вернемся после того как рассмотрим материал, относящийся к экспериментальным данным, полученным на амфибиях.
В опытах с тритонами (см. главу восьмую «Амфибии») для образования близнецов путем перетяжки яйца Шпеман (Spemann u. Falkenberg, 1919) попутно получил интересный материал для суждения о возникновении s. i. у этих животных. Из 36 пар свободных ОБ, полученных в большинстве благодаря перетяжке бластулы, из 25 левых партнеров 24 имели нормальное расположение внутренностей и лишь у одного было правосердие, т. е. частичный s. i. Из 30 правых близнецов у 15 сердце и кишечник имели нормальное расположение, но у 14 был s. i., у одного — неясная картина. Иначе говоря, почти у половины правых партнеров был s. i., как и у рыб по данным Комаи и Линна. У свободных близнецов иногда имелось искривление тела с той стороны, с которой они ранее были соединены со своим близнецом, т. е. правый близнец загибался налево (рис.59),
особенно сделанные сравнительно недавно — Комаи (Komai, 1938) и Линна (Lynn, 1946), работавших на лососевых разных видов. Комаи изучил s. i. у 502 пар «свободных» ОБ, связанных только желточным мешком, и у 287 пар двойных уродств, в большинстве с передним раздвоением. У «свободных» близнецов он нашел 17.5% особен с s. i. при 6.5% у одиночек. У деформированных особей свободных близнецов s. i. наблюдался в 3 раза чаще, чем у нормальных, т. е. обнаружена корреляция между деформацией и s. i. У соединенных близнецов правый компонент был чаще деформирован и недоразвит, чем левый, и s. i. правых близнецов оказался у 66 пар, тогда как s. i. левых близнецов был только у 31 пары, т. е. в 2 раза реже.
С еще большим материалом работал Линн. Он изучил 2100 близнецов (свободных и соединенных) и 1110 одиночных особей, обращая внимание на положение и форму желудка, печени и плавательного пузыря. Одиночные особи в материале Линна имели s. i. в 4.7%. Больший процент (29.5) с s. i. был среди особей с искривлением тела в одну сторону. Интересно, что s. i. чаще встречается у особей с таким искривлением тела влево (44.7%), чем вправо (14.3%).
Среди близнецов s. i. встречался значительно чаще, чем среди одиночных особей — 37.2%. Соединенные близнецы имели s. i. чаще (41.1%), чем свободные (30.8%). В группе асимметричных соединенных близнецов, где один из партнеров оказывается недоразвитым или дефективным, паразитом, s. i. чаще встречается именно у такого близнеца, а не у более развитого (автозита). Интересно, что и по данным Линна правый близнец у соединенных имеет s. i. вдвое чаще, чем левый (22.3 % к 11.5%). Иное отношение, однако, найдено у швейцарской форели (Bovet, 1931), правда, на сравнительно небольшом материале (42 пары): у правых компонентов s. i. обнаружен в меньшем числе, чем у левых (на 9 правых компонентов 15 левых). Единичные случаи s. i., у левого близнеца описаны также у амфибий (Schwind, 1942).
Интересно, что встречаются такие случаи, когда оба свободных близнеца имеют s. i. Среди соединенных близнецов это явление наблюдается чаще (8% по Бове и 7.3% по Линну). Место соединения также влияет на частоту s. i. Чаще всего он встречается у близнецов, соединенных на уровне спинного плавника, т. е. приблизительно посередине тела.
Линн (Lynn a. Peadon, 1949) изучал и троен тех же рыб (рис.55).
Оказалось, что s. i. у одного и более компонентов тройни встречается еще чаще, чем у двоен, а именно в количестве 70.9%.
Б общем, очевидно, существует тесная зависимость между процессом образования близнецов, возникновением дефектов развития и зеркальностью внутренних органов (s. i.). Физиологический механизм этого явления еще мало выяснен. К существующим теоретическим интерпретациям зеркальности близнецов мы еще вернемся после того как рассмотрим материал, относящийся к экспериментальным данным, полученным на амфибиях.
В опытах с тритонами (см. главу восьмую «Амфибии») для образования близнецов путем перетяжки яйца Шпеман (Spemann u. Falkenberg, 1919) попутно получил интересный материал для суждения о возникновении s. i. у этих животных. Из 36 пар свободных ОБ, полученных в большинстве благодаря перетяжке бластулы, из 25 левых партнеров 24 имели нормальное расположение внутренностей и лишь у одного было правосердие, т. е. частичный s. i. Из 30 правых близнецов у 15 сердце и кишечник имели нормальное расположение, но у 14 был s. i., у одного — неясная картина. Иначе говоря, почти у половины правых партнеров был s. i., как и у рыб по данным Комаи и Линна. У свободных близнецов иногда имелось искривление тела с той стороны, с которой они ранее были соединены со своим близнецом, т. е. правый близнец загибался налево (рис.59),
 а левый направо. Такую асимметрию Шпеман объяснял недостатком материала на той стороне, с которой отделился другой близнец и где должен был восстанавливаться утраченный этим путем материал. В этих случаях асимметрии правый близнец иногда обнаруживал s. i. Эта связь у тритонов оказалась мало изученной. По видимому, эти искривленные тритоны аналогичны описанным Линном искривленным рыбам.
Опыты Шпемана с амфибиями показали, что степень асимметрии близнецов зависит от того, на какой стадии развития накладывается лигатура. Перетяжка на стадии двух или четырех бластомеров лишь иногда имела следствием появление слабо выраженной асимметрии, тогда как перетяжка на стадии гаструлы весьма значительную, т. е. регуляция была тем менее совершенна, чем позже произошло раздвоение зародыша.
У соединенных близнецов тритонов с раздвоенным передним концом s. i. обнаружен у большинства правых компонентов (рис.60) .
а левый направо. Такую асимметрию Шпеман объяснял недостатком материала на той стороне, с которой отделился другой близнец и где должен был восстанавливаться утраченный этим путем материал. В этих случаях асимметрии правый близнец иногда обнаруживал s. i. Эта связь у тритонов оказалась мало изученной. По видимому, эти искривленные тритоны аналогичны описанным Линном искривленным рыбам.
Опыты Шпемана с амфибиями показали, что степень асимметрии близнецов зависит от того, на какой стадии развития накладывается лигатура. Перетяжка на стадии двух или четырех бластомеров лишь иногда имела следствием появление слабо выраженной асимметрии, тогда как перетяжка на стадии гаструлы весьма значительную, т. е. регуляция была тем менее совершенна, чем позже произошло раздвоение зародыша.
У соединенных близнецов тритонов с раздвоенным передним концом s. i. обнаружен у большинства правых компонентов (рис.60) .
 Шпеманом, его учениками и последователями было проделано несколько экспериментальных работ с целью искусственного вызывания s. i. у тритонов и объяснения причин этого явления. Данные одной из последних работ (Woellwarth, 1950) проливают некоторый свет на связь s. i. с близнецами. Вёллворт на большом материале повторял и дополнил один из основных опытов школы Шпемана (Spemann, 1906; Pressler, 1911; Meyer, 1932), согласно которому четырехугольная пластинка тканей, вырезанная из середины нейрулы тритона и после поворота на 180° посаженная обратно, вызывает в некотором числе случаев полный s. i. Толкование этого опыта было спорно и неясно. Вёллворт попытался это явление объяснить той же причиной, какой вызывается s. i. у тритонов-близнецов с изгибом тела, о которых речь была выше: а именно, дефектом, изъяном тканей, утраченных при отделении другого близнеца. В опытах с вырезанием пластинки из нейрулы также происходит частичная утрата тканей, вызывающая у части животных s. i. Правильность этой гипотезы («Defekthypothese») автор показал на опытах с простым удалением кусочков нейрулы без всяких поворотов вырезанного участка. Особенно демонстративно обнаружилось влияние нанесения дефекта на стадии гаструлы в зависимости от стороны, на которую дефект наносился: если слева, то получалось 56% инверсии, а если справа, то только 8%. Это объясняется так: дефект, нехватка тканей вызывает изгиб кишечника в сторону дефекта. Если этот изгиб, вызванный дефектом, совпадает с естественным изгибом, соответствующим нормальной асимметрии (рис.61), то таковая и сохраняется. Это бывает при дефекте справа. Когда же дефект нанесен слева, то вызванное им искривление оказывается в антагонизме с естественным искривлением, и если тенденция, вызванная дефектом, берет верх, то возникает s. i. Этим объясняется совпадение большего числа случаев s. i. у одиночных особей с дефектом, нанесенным слева, и правых искривленных свободных близнецов тритонов и рыб, ибо отделение их левого близнеца нанесло им дефект тоже слева. Интересно, что в естественной популяции тритонов (просмотрено 659 особей) s. i. оказался у 5, т. е. у 0.76%.
Шпеманом, его учениками и последователями было проделано несколько экспериментальных работ с целью искусственного вызывания s. i. у тритонов и объяснения причин этого явления. Данные одной из последних работ (Woellwarth, 1950) проливают некоторый свет на связь s. i. с близнецами. Вёллворт на большом материале повторял и дополнил один из основных опытов школы Шпемана (Spemann, 1906; Pressler, 1911; Meyer, 1932), согласно которому четырехугольная пластинка тканей, вырезанная из середины нейрулы тритона и после поворота на 180° посаженная обратно, вызывает в некотором числе случаев полный s. i. Толкование этого опыта было спорно и неясно. Вёллворт попытался это явление объяснить той же причиной, какой вызывается s. i. у тритонов-близнецов с изгибом тела, о которых речь была выше: а именно, дефектом, изъяном тканей, утраченных при отделении другого близнеца. В опытах с вырезанием пластинки из нейрулы также происходит частичная утрата тканей, вызывающая у части животных s. i. Правильность этой гипотезы («Defekthypothese») автор показал на опытах с простым удалением кусочков нейрулы без всяких поворотов вырезанного участка. Особенно демонстративно обнаружилось влияние нанесения дефекта на стадии гаструлы в зависимости от стороны, на которую дефект наносился: если слева, то получалось 56% инверсии, а если справа, то только 8%. Это объясняется так: дефект, нехватка тканей вызывает изгиб кишечника в сторону дефекта. Если этот изгиб, вызванный дефектом, совпадает с естественным изгибом, соответствующим нормальной асимметрии (рис.61), то таковая и сохраняется. Это бывает при дефекте справа. Когда же дефект нанесен слева, то вызванное им искривление оказывается в антагонизме с естественным искривлением, и если тенденция, вызванная дефектом, берет верх, то возникает s. i. Этим объясняется совпадение большего числа случаев s. i. у одиночных особей с дефектом, нанесенным слева, и правых искривленных свободных близнецов тритонов и рыб, ибо отделение их левого близнеца нанесло им дефект тоже слева. Интересно, что в естественной популяции тритонов (просмотрено 659 особей) s. i. оказался у 5, т. е. у 0.76%.
 Вёллворт изучил связь (корреляцию) s. i. с одним асимметричным парным участком промежуточного мозга, nuclei rabenulae, где левое ядро этого участка крупнее и по структуре отличается от правого. И хотя при всех операциях этот участок мозга, находясь на значительном отдалении, никогда не затрагивался, он и при возникновении s. i. также оказывался инвертированным, обнаруживая корреляцию, близкую к таковой между кишечником и сердцем (около 80%). Физиологический механизм этой зависимости пока остается неясным.
«Теория дефекта» Вёллворта, объясняя возникновение s. i. и его преобладание у правых близнецов, объясняет и связь s. i. с различными аномалиями. Последние вызваны тем же «дефектом», которым вызван s. i. Новые исследования покажут, насколько эта теория верна и применима к другим животным, в частности к иглокожим, на которых получены некоторые данные, аналогичные описанным у тритонов (Newman, 1923, и др.).
Изложенная теория возникновения s. i. предполагает, что существует какой-то физиологический механизм, благодаря которому осуществляется нормальная асимметрия, инверсией которой является s. i. Способ возникновения этой нормальной асимметрии пока остается невыясненным. Некоторые исследователи предполагают, что в связи с возникновением главного градиента активности возникает также билатеральная асимметрия, и левая сторона животного обладает большей физиологической активностью, чем правая, что и подтверждается некоторыми опытами (Гексли и де-Бер, 1936). Соображениями о большей активности левой стороны пытаются также подойти к объяснению разной частоты s. i. у правых и левых близнецов, а именно: при разрезании развивающегося яйца пополам поверхность разреза повреждается; от этого в левой половине градиент асимметрии не меняется, т. е. левая сторона остается активнее новой правой. В правой же половине могут произойти изменения: эта сторона в данном случае окажется активнее новой левой (если не произойдет регуляции), тогда возникнет обращенная асимметрия и может появиться s. i. Эти соображения, однако, еще мало обоснованы фактами.
Недавно была сделана попытка получить близнецов у тритонов путем рассечения ранней гаструлы на правую и левую половины (Mangold u. Testa, 1953; Mangold et al., 1956). При такой операции каждая полугаструла остается с половинной массой зачатков разных органов и резко нарушенной симметрией. В одном из вариантов этих опытов только ткань будущего эпидермиса, снятая с третьей гаструлы, приращивалась к опытным полугаструлам, благодаря чему эта ткань оказывалась в избыточной массе. При этом варианте опыта наблюдалась, как правило, далеко идущая регуляция. Парные органы развиваются, и в некоторых случаях наблюдается лишь незначительное недоразвитие тех из них, которые находятся на той стороне эмбриона, с которой происходило разделение: с левой стороны у зародыша из правой полугаструлы, с правой — у левого зародыша. Но наряду с регуляцией наблюдался также различные недостатки развития и возникновение разных аномалий в зависимости от многих условий протекания операции и последующего развития.
У 7 пар таких искусственных близнецов изучалось развитие внутренних органов на гистологических срезах. Регуляция разных органов происходила неодинаково. Так, например, средний мозг в большей мере, чем другие органы, достигал полной симметрии. В разной степени отставали от него в этом отношении различные другие органы. Низшую степень регуляции обнаруживали, например, миотомы: они оказались заметно несимметричными и не только передние, но также и хвостовые, хотя последние развиваются позже передних. Из 7 пар близнецов у 6 правых близнецов удалось установить s. i. внутренних органов, у левых - только у одного и то неполный s. i., а только лишь сердца.
Зеркальная асимметрия проявляется также в ряде случаев, установленных экспериментально, когда две особи или две системы органов развиваются бок о бок, связанные участком тканей друг с другом. Они образуют одну систему из двух и более взаимновлияющих друг на друга компонентов, разделенных плоскостями симметрии. Механизм этого влияния не выяснен. Может быть, отношения, возникающие в связи с повреждением, согласно теории Вёллворта, надо рассматривать лишь как добавочные на этой общей основе. Примером, аналогичным соединенным близнецам, могут служить наблюдения и опыты с конечностями амфибий — здесь один компонент как бы индуцирует симметричный себе градиент, вследствие чего получается зеркальная асимметрия другого (Child, 1941).
Добавочные конечности у эмбрионов амфибии возникают под влиянием различных повреждений почки конечности, в частности пересадки рядом с ней почки другой конечности с соблюдением известных условий (Mangold, 1929). Зеркальность добавочных конечностей (вторичной и третичной) отчетливо выступает в таких случаях, причем основные оси этих конечностей лежат в одной плоскости, а зеркальная плоскость между двумя конечностями делит пополам угол, под которым они лежат но отношению друг к другу (рис. 62).
Вёллворт изучил связь (корреляцию) s. i. с одним асимметричным парным участком промежуточного мозга, nuclei rabenulae, где левое ядро этого участка крупнее и по структуре отличается от правого. И хотя при всех операциях этот участок мозга, находясь на значительном отдалении, никогда не затрагивался, он и при возникновении s. i. также оказывался инвертированным, обнаруживая корреляцию, близкую к таковой между кишечником и сердцем (около 80%). Физиологический механизм этой зависимости пока остается неясным.
«Теория дефекта» Вёллворта, объясняя возникновение s. i. и его преобладание у правых близнецов, объясняет и связь s. i. с различными аномалиями. Последние вызваны тем же «дефектом», которым вызван s. i. Новые исследования покажут, насколько эта теория верна и применима к другим животным, в частности к иглокожим, на которых получены некоторые данные, аналогичные описанным у тритонов (Newman, 1923, и др.).
Изложенная теория возникновения s. i. предполагает, что существует какой-то физиологический механизм, благодаря которому осуществляется нормальная асимметрия, инверсией которой является s. i. Способ возникновения этой нормальной асимметрии пока остается невыясненным. Некоторые исследователи предполагают, что в связи с возникновением главного градиента активности возникает также билатеральная асимметрия, и левая сторона животного обладает большей физиологической активностью, чем правая, что и подтверждается некоторыми опытами (Гексли и де-Бер, 1936). Соображениями о большей активности левой стороны пытаются также подойти к объяснению разной частоты s. i. у правых и левых близнецов, а именно: при разрезании развивающегося яйца пополам поверхность разреза повреждается; от этого в левой половине градиент асимметрии не меняется, т. е. левая сторона остается активнее новой правой. В правой же половине могут произойти изменения: эта сторона в данном случае окажется активнее новой левой (если не произойдет регуляции), тогда возникнет обращенная асимметрия и может появиться s. i. Эти соображения, однако, еще мало обоснованы фактами.
Недавно была сделана попытка получить близнецов у тритонов путем рассечения ранней гаструлы на правую и левую половины (Mangold u. Testa, 1953; Mangold et al., 1956). При такой операции каждая полугаструла остается с половинной массой зачатков разных органов и резко нарушенной симметрией. В одном из вариантов этих опытов только ткань будущего эпидермиса, снятая с третьей гаструлы, приращивалась к опытным полугаструлам, благодаря чему эта ткань оказывалась в избыточной массе. При этом варианте опыта наблюдалась, как правило, далеко идущая регуляция. Парные органы развиваются, и в некоторых случаях наблюдается лишь незначительное недоразвитие тех из них, которые находятся на той стороне эмбриона, с которой происходило разделение: с левой стороны у зародыша из правой полугаструлы, с правой — у левого зародыша. Но наряду с регуляцией наблюдался также различные недостатки развития и возникновение разных аномалий в зависимости от многих условий протекания операции и последующего развития.
У 7 пар таких искусственных близнецов изучалось развитие внутренних органов на гистологических срезах. Регуляция разных органов происходила неодинаково. Так, например, средний мозг в большей мере, чем другие органы, достигал полной симметрии. В разной степени отставали от него в этом отношении различные другие органы. Низшую степень регуляции обнаруживали, например, миотомы: они оказались заметно несимметричными и не только передние, но также и хвостовые, хотя последние развиваются позже передних. Из 7 пар близнецов у 6 правых близнецов удалось установить s. i. внутренних органов, у левых - только у одного и то неполный s. i., а только лишь сердца.
Зеркальная асимметрия проявляется также в ряде случаев, установленных экспериментально, когда две особи или две системы органов развиваются бок о бок, связанные участком тканей друг с другом. Они образуют одну систему из двух и более взаимновлияющих друг на друга компонентов, разделенных плоскостями симметрии. Механизм этого влияния не выяснен. Может быть, отношения, возникающие в связи с повреждением, согласно теории Вёллворта, надо рассматривать лишь как добавочные на этой общей основе. Примером, аналогичным соединенным близнецам, могут служить наблюдения и опыты с конечностями амфибий — здесь один компонент как бы индуцирует симметричный себе градиент, вследствие чего получается зеркальная асимметрия другого (Child, 1941).
Добавочные конечности у эмбрионов амфибии возникают под влиянием различных повреждений почки конечности, в частности пересадки рядом с ней почки другой конечности с соблюдением известных условий (Mangold, 1929). Зеркальность добавочных конечностей (вторичной и третичной) отчетливо выступает в таких случаях, причем основные оси этих конечностей лежат в одной плоскости, а зеркальная плоскость между двумя конечностями делит пополам угол, под которым они лежат но отношению друг к другу (рис. 62).
 И хотя физиологический механизм данных явлений на амфибиях еще не изучен, все же экспериментальные исследования на этих животных дали уже многое для понимания феномена зеркальности.
У человеческих близнецов s. i. описан как большая редкость. Возможно, что в ряде случаев, особенно при абортах и мертворожденных s. i. не регистрировался и потому мог оказаться незамеченным. Ньюмен (Newman, 1940b) писал, что известно только 5 пар ОБ, где в каждой паре один из близнецов имеет s. i. С тех пор, по-видимому, обнаружен еще только один случай (Jeune et Confavreux, 1948). Вероятно, еще более редко возникают такие ОБ, когда у обоих близнецов имеется s. i. Пока что в литературе известно 3 таких пары (Reinhardt, 1913; Pezzi е Carugati, 1924; Kean, 1942). Последняя из этих пар — 18-летние девушки — была довольно подробно изучена. Рентгенограммы показали полный s. i. у обеих. Электрокардиограммы у них оказались инвертированными. Обе близнячки были правшами, но неизвестно, с детства ли. Родились они с разными плацентами и при рождении имели разный вес. Наконец, надо отметить, что описан один случай РБ (Torgersen, 1948), где оба близнеца имели s. i., причем у одного он был полный, а у другого только в брюшной области. Как могли возникнуть такие разнояйцевые близнецы, оба имеющие s. i., остается пока что непонятным.
Гораздо чаще, чем среди свободных ОБ, встречается s. i. у соединенных близнецов, где это скорее правило, а отсутствие s. i. у одного из компонентов - исключение (рис. 58, 63; Newman, 1923, 1931, 1940b). Правый близнец имеет иногда дефекты, которых нет у левого, как например недоразвитие матки у Иозефы Блажек. У правого же компонента чаще встречается s. i., хотя таковой описан иногда и у левого. К сожалению, относительно существования s. i. у «сиамских близнецов» разных вариантов, живших в конце XIX в. и в начале XX в. (Чанг-Энг и др.), почти ничего неизвестно.
И хотя физиологический механизм данных явлений на амфибиях еще не изучен, все же экспериментальные исследования на этих животных дали уже многое для понимания феномена зеркальности.
У человеческих близнецов s. i. описан как большая редкость. Возможно, что в ряде случаев, особенно при абортах и мертворожденных s. i. не регистрировался и потому мог оказаться незамеченным. Ньюмен (Newman, 1940b) писал, что известно только 5 пар ОБ, где в каждой паре один из близнецов имеет s. i. С тех пор, по-видимому, обнаружен еще только один случай (Jeune et Confavreux, 1948). Вероятно, еще более редко возникают такие ОБ, когда у обоих близнецов имеется s. i. Пока что в литературе известно 3 таких пары (Reinhardt, 1913; Pezzi е Carugati, 1924; Kean, 1942). Последняя из этих пар — 18-летние девушки — была довольно подробно изучена. Рентгенограммы показали полный s. i. у обеих. Электрокардиограммы у них оказались инвертированными. Обе близнячки были правшами, но неизвестно, с детства ли. Родились они с разными плацентами и при рождении имели разный вес. Наконец, надо отметить, что описан один случай РБ (Torgersen, 1948), где оба близнеца имели s. i., причем у одного он был полный, а у другого только в брюшной области. Как могли возникнуть такие разнояйцевые близнецы, оба имеющие s. i., остается пока что непонятным.
Гораздо чаще, чем среди свободных ОБ, встречается s. i. у соединенных близнецов, где это скорее правило, а отсутствие s. i. у одного из компонентов - исключение (рис. 58, 63; Newman, 1923, 1931, 1940b). Правый близнец имеет иногда дефекты, которых нет у левого, как например недоразвитие матки у Иозефы Блажек. У правого же компонента чаще встречается s. i., хотя таковой описан иногда и у левого. К сожалению, относительно существования s. i. у «сиамских близнецов» разных вариантов, живших в конце XIX в. и в начале XX в. (Чанг-Энг и др.), почти ничего неизвестно.

Пока, по-видимому, неизвестно ни одного случая, когда бы оба соединенные близнеца имели s. i. (Newman, 1940b). Между данными относительно s. i. у людей-близнецов и близнецов различных животных имеется ясная аналогия. Мы рассмотрели ряд асимметричных признаков, которые у близнецов могут иметь зеркальный характер. Можно поставить вопрос: существует ли какая-нибудь зависимость между возникновением близнецов и наличием у них зеркальности ряда асимметричных признаков? На этот вопрос пытаются ответить некоторые гипотезы. Мы рассмотрим две из них, существенно отличающиеся друг от друга: одна приписывает возникновение внутрипарной зеркальности у ОБ их возникновению под влиянием внешних факторов, другая — генотипическим различиям. Первую гипотезу в течение ряда лет развивал и обосновал главным образом Ньюмен (Newman, 1923—1940). Сущность ее заключается в том, что зеркальность ОБ есть не что иное, как проявление зеркальности правой и левой стороны того первоначального единого зародыша, от продольного раздвоения которого произошли ОБ. Ньюмен не вдается в рассмотрение исходного различия правой и левой стороны зародыша, соглашаясь с представлением о существовании латерального градиента и большей физиологической активности левой стороны. Как мы уже знаем, Ньюмен вместе со Стоккардом (Stockard, 1921) считают, что факторы среды, задерживающие развитие зародыша, являются причиной возникновения ОБ, а в связи с этим и их зеркальных различий. Чем же можно объяснить, что у большинства ОБ зеркальность отсутствует и почему одни асимметрические структуры иногда обнаруживают зеркальность, в то время как другие ее не проявляют? Ньюмен считает, что это зависит от стадии развития зародыша, на которой произошло раздвоение. Разумеется, что чем раньше произошло раздвоение, тем полнее должна быть регуляция обеих половин. Надо думать, что на ранних стадиях развития зародыша вообще еще отсутствует или очень лабильно заложена дифференцировка на правую и левую сторону, почему и нельзя ожидать проявления зеркальности при возникновении ОБ на ранних стадиях эмбрионального развития. Такое предположение правдоподобно, но оно еще мало обосновано фактами. Вообще зеркальность, по Ньюмену, зависит от трех факторов:
а) времени раздвоения, б) от степени зависимости разных асимметрий от разных моментов развития в) степени возможной регуляции в смысле восстановления нормы.В пользу гипотезы Ньюмена говорит, во всяком случае, тот факт, что у соединенных близнецов, которых считают произошедшими на сравнительно поздних стадиях развития, зеркальность и, в частности, s. i. выражены гораздо значительнее и чаще, чем у свободных ОБ. В пользу своей гипотезы Ньюмен приводит многочисленные факты как из своих наблюдений и опытов, так и почерпнутые из работ других ученых (вышеприведенные данные Комаи и др.). В частности, Ньюмен ссылается на небольшую статью Килера (Keeler, 1929), в которой делается сравнение степени зеркальности у свободных и соединенных человеческих ОБ. Для сравнения служили следующие четыре признака: отпечатки узоров кожи рук, неправильности зубов, завиток волос на темени и форма ушей. Сравнивались 14 свободных ОБ и 10 пар соединенных. Последние делились на две группы: соединенные латерально — 6 пар и соединенные вентрально — 4 пары. У свободных близнецов обнаружено 5 случаев зеркальности (22%) на 18 случаев (78%) отсутствия таковой. У 6 пар соединенных латерально близнецов оказалось 10 случаев зеркальных признаков (77%) и 3 случая (23%) отсутствия зеркальности. У соединенных вентрально лишь у одной пары и только один признак (завиток на темени) был зеркальным (13%), а остальные одинаковы (87%). Различие между свободными и латерально соединенными близнецами в смысле зеркальности в этом материале Килера выступает довольно отчетливо. Интересно отметить, что у вентрально соединенных близнецов зеркальность почти отсутствует. В материале Килера ничего, к сожалению, не говорится о s. i. 6 пар, соединенных латерально, у которых обнаружено 10 случаев зеркальности мелких асимметричных признаков. Однако Ньюмен (Newman, 1940b) сопоставляет избыток зеркальности мелких признаков у соединенных близнецов в материале Килера с избытком s. i. у соединенных близнецов рыб по Комаи и усматривает на основании этого корреляцию между s. i. и другими случаями асимметрии. Он делает вывод, что последние признаки суть проявление подлинной зеркальности ОБ. Сопоставление столь разного, а у Килера и небольшого материала, вызывает сомнение в правильности этого вывода и для обоснования его необходим новый, более убедительный материал, чем собранный Килером. К сожалению, с 1929 г., когда вышла статья Килера, до сих пор ни сам Ньюмен, ни другие исследователи никаких новых доказательств, насколько мне известно, не представили. Вопреки Ньюмену, корреляция между различными асимметрическими признаками, оказывающимися иногда зеркальными у ОБ, не может считаться доказанной, тем более, что против нее и вообще против зеркальности таких признаков было приведено в литературе довольно много фактов. Что касается корреляции между разными из упомянутых признаков, то ею занимались некоторые ученые. Фершюр (Verschuer, 1932) вычислял корреляцию между завитком волос на темени и леворукостью, манерой складывать руки и другими, но никакой корреляции не обнаружил. Это признаки далекие, а такой признак, как манера складывать руки, вообще вряд ли имеет прямое отношение к прирожденным свойствам человека. Райф (Rife, 1933b) у 20 пар ОБ сопоставил три признака: леворукость, преобладание одного глаза над другим и завиток на темени. Эти признаки оказались в зеркальном взаимоотношении по-разному: у 15 пар обнаружилось преобладание глаза, частичное или полное; у 8 пар один из партнеров был левша, тоже частично или полностью; наконец, 6 пар имели различие по теменному завитку волос при наличии 5 пар, у которых картина была неясной. Между леворукостью и преобладанием левого глаза установлено полное совпадение, что и можно ожидать ввиду их функциональных связей и зависимости от одного полушария мозга. Обратный же ход завитка волос, по-видимому, независим от двух других признаков. Другая группа фактов относится к проверке ожидаемого теорией Ньюмена большего количества случаев зеркальности между разными асимметричными признаками у ОБ, чем и массе населения. Приведенные выше данные относительно численности левшей среди близнецов показывают, что зеркальность выражена приблизительно одинаково как среди РБ, так и ОБ. Это не говорит в пользу теории Ньюмена относительно этого признака. Фершюр (Verschuer, 1932) изучил распределение некоторых признаков, например завитка на темени, у 163 пар ОБ и сопоставил эмпирические цифры с цифрами расчета «случайного» распределения (табл. 8).
 На основании своих данных Фершюр возражает против гипотезы Ньюмена в отношении свободных ОБ и считает, что наблюдаемые у них явления зеркальности — результат случайного совпадения, не имеют прямого отношения к происхождению близнецов и вызваны они факторами пре- или постнатальной среды. Он допускает исключение лишь для некоторых случаев ОБ, которые возникли на поздней ступени эмбрионального развития, когда между правой и левой половиной уже сложились известные различия, которые ясно выступают у соединенных близнецов.
Комаи и Фукуока (Komai a. Fukuoka, 1943b) на основании своего материала также скептически относятся к зеркальности у ОБ этих двух признаков (леворукости и теменного завитка).
Аналогичные соображения высказаны Райфом и Камменсом (Rife a. Cummins, 1943) относительно отпечатков кожных узоров ладоней ОБ, на основании их сравнения с РБ, братьями-сестрами и популяцией: зеркальность дерматоглифов у ОБ встречается не чаще, чем у любой пары людей. Связь между леворукостью, s. i. и возникновением близнецов отрицает также Торгерсен (Torgersen, 1950) и другие.
В защиту своих взглядов Ньюмен (Newman, 1940b) мог только возразить, что его противники путают «псевдозеркальность», наблюдаемую у РБ и в популяции с «истинной» зеркальностью ОБ. Никаких доказательств существования двух таких типов зеркальности Ньюмен не приводит.
Таково положение с вопросом о зеркальности свободных ОБ. Он, как видно, не решен окончательно, но в общем факты говорят скорее против гипотезы Ньюмена, чем в пользу нее. Иначе обстоит дело с соединенными близнецами. Ньюмен считает, что здесь s. i. у одного из компонентов является скорее правилом, чем исключением. Это положение находит отчасти подтверждение в рассмотренном уже материале соединенных близнецов у рыб. К сожалению, оно еще мало подтверждено на высших животных, например домашних, и на человеке. Сам Ньюмен (Newman, 1931), описывая ныне живущих или недавно живших соединенных человеческих близнецов (Блажек, Хилтон, Родена и др.), ничего не говорит относительно s. i. у кого-нибудь из них. Также нет сведений о s. i. у соединенных близнецов домашних животных (Keller и. Niedoba, 1937, и др.). Что касается других признаков, то они, по-видимому, чаще обнаруживают зеркальность, как уже выше говорилось на основании работы Килера и других авторов, в частности, относительно кожных узоров (Newman, 1931, и др.). Кроме прямых признаков зеркальности, соединенные близнецы в ряде случаев обнаруживают заметные внутрипарные различия, в результате которых соединенные партнеры оказываются более непохожими, чем свободные партнеры ОБ. Это явление, на первый взгляд парадоксальное, так как соединенные симметричные близнецы менее всего могут вызывать сомнение в своей однояйцевости, Ньюмен объясняет тем, что правый и левый компоненты в соединенной паре в большей мере проявляют различия, вызванные их происхождением из одной из половин зародыша, чем свободные ОБ (Newman, 1931). Сюда относятся и различные дефекты и аномалии развития, которые описаны также и у домашних животных (Keller и. Niedoba, 1937). В этом вопросе Ньюмену по существу не возражает и Фершюр (Verschuer, 1932). Он справедливо считает, что ОБ надо изучать в связи со стадией развития, на которой произошло раздвоение, и соответственно стадией закладки того или иного органа или признака. Зеркальность асимметричных признаков не может иногда не зависеть от стадии развития, на которой произошло раздвоение (Verschuer, 1929). К сожалению, этот вопрос до сих пор еще очень мало разработан.
Таким образом, по отношению к соединенным ОБ гипотеза Ньюмена кажется несколько более обоснованной, чем по отношению к свободным ОБ. Однако достаточно доказанной ее все же считать нельзя, и для полного выяснения правильности её или степени правильности необходимы дальнейшие исследования. Во всяком случае, в качестве рабочей гипотезы она была полезна и остается таковой и в настоящее время. Многие вопросы эта гипотеза до сих пор не может объяснить. Сам Ньюмен (Newman, 1940b) указал некоторые из них. Почему среди ОБ могут чаще встречаться признаки зеркальности, чем среди одиночек? Почему у ОБ одни признаки оказываются зеркальными, а другие нет — например при s. i. у одного из близнецов теменной завиток у него идет нормально? Почему у одних свободных ОБ с одинаковой степенью сходства зеркальность выступает резче, а у других слабее? Почему у одинаково соединенных близнецов может быть разное выражение зеркальности? Нет надобности увеличивать число таких вопросов; приведенных достаточно, чтобы показать, насколько вопрос о зеркальности у близнецов еще неясен, как много еще непонятного в нем.
Другого рода гипотезы были высказаны для объяснения зеркальности асимметричных признаков на основе генетики. В области наследственности асимметрии есть несколько интересных исследований, например Пшибрама (Phzibram, 1908) о наследственности разного цвета правого и левого глаза у ангорских кошек или Бойкотта с сотрудниками (Boycott et al., 1929) над улитками, имеющими спирально завитую раковину направо или налево. Объяснение, даваемое этим явлениям, сводилось на менделирование определенных генов. Дальберг (Dahlberg, 1926, 1929, 1943/44) по аналогии с этими явлениями высказал предположение о существовании неодинакового распределения генов при образовании ОБ, отчего зависит зеркальность асимметричных признаков, назвав свою гипотезу «теорией генотипической асимметрии». Эта гипотеза носит умозрительный характер и недостаточно подтверждается фактами. Несколько иначе, но в том же направлении Боутервек (Bouterwek, 1943) старался доказать, что асимметрия и зеркальность ОБ основаны на различии их генотипов. Этот тезис и его доказательства вызвали ряд возражений (Lotze, 1937, и др.), и действительно, попытки Боутервека не производят убедительного впечатления. Вопрос о значении генотипа для зеркальности у близнецов остается пока что нерешенным. Генотип, вероятно, играет известную роль и, может быть, неодинаковую у разных групп животных (Torgersen, 1950). Однако понять роль генотипа можно будет только тогда, когда глубже будет изучен физиологический механизм становления асимметрии зародыша, изменение этой асимметрии в зависимости от раздвоения зародыша и в связи с этим возможности возникновения зеркальности на разных ступенях развития эмбриона. Тогда будет также изжит односторонний характер гипотезы Ньюмена.
Теперь, после рассмотрения некоторых общих вопросов близнецовой проблемы, мы можем перейти к знакомству с многообразным проявлением близнечества в различных классах животного царства, начиная с низших форм и кончая человеком.
На основании своих данных Фершюр возражает против гипотезы Ньюмена в отношении свободных ОБ и считает, что наблюдаемые у них явления зеркальности — результат случайного совпадения, не имеют прямого отношения к происхождению близнецов и вызваны они факторами пре- или постнатальной среды. Он допускает исключение лишь для некоторых случаев ОБ, которые возникли на поздней ступени эмбрионального развития, когда между правой и левой половиной уже сложились известные различия, которые ясно выступают у соединенных близнецов.
Комаи и Фукуока (Komai a. Fukuoka, 1943b) на основании своего материала также скептически относятся к зеркальности у ОБ этих двух признаков (леворукости и теменного завитка).
Аналогичные соображения высказаны Райфом и Камменсом (Rife a. Cummins, 1943) относительно отпечатков кожных узоров ладоней ОБ, на основании их сравнения с РБ, братьями-сестрами и популяцией: зеркальность дерматоглифов у ОБ встречается не чаще, чем у любой пары людей. Связь между леворукостью, s. i. и возникновением близнецов отрицает также Торгерсен (Torgersen, 1950) и другие.
В защиту своих взглядов Ньюмен (Newman, 1940b) мог только возразить, что его противники путают «псевдозеркальность», наблюдаемую у РБ и в популяции с «истинной» зеркальностью ОБ. Никаких доказательств существования двух таких типов зеркальности Ньюмен не приводит.
Таково положение с вопросом о зеркальности свободных ОБ. Он, как видно, не решен окончательно, но в общем факты говорят скорее против гипотезы Ньюмена, чем в пользу нее. Иначе обстоит дело с соединенными близнецами. Ньюмен считает, что здесь s. i. у одного из компонентов является скорее правилом, чем исключением. Это положение находит отчасти подтверждение в рассмотренном уже материале соединенных близнецов у рыб. К сожалению, оно еще мало подтверждено на высших животных, например домашних, и на человеке. Сам Ньюмен (Newman, 1931), описывая ныне живущих или недавно живших соединенных человеческих близнецов (Блажек, Хилтон, Родена и др.), ничего не говорит относительно s. i. у кого-нибудь из них. Также нет сведений о s. i. у соединенных близнецов домашних животных (Keller и. Niedoba, 1937, и др.). Что касается других признаков, то они, по-видимому, чаще обнаруживают зеркальность, как уже выше говорилось на основании работы Килера и других авторов, в частности, относительно кожных узоров (Newman, 1931, и др.). Кроме прямых признаков зеркальности, соединенные близнецы в ряде случаев обнаруживают заметные внутрипарные различия, в результате которых соединенные партнеры оказываются более непохожими, чем свободные партнеры ОБ. Это явление, на первый взгляд парадоксальное, так как соединенные симметричные близнецы менее всего могут вызывать сомнение в своей однояйцевости, Ньюмен объясняет тем, что правый и левый компоненты в соединенной паре в большей мере проявляют различия, вызванные их происхождением из одной из половин зародыша, чем свободные ОБ (Newman, 1931). Сюда относятся и различные дефекты и аномалии развития, которые описаны также и у домашних животных (Keller и. Niedoba, 1937). В этом вопросе Ньюмену по существу не возражает и Фершюр (Verschuer, 1932). Он справедливо считает, что ОБ надо изучать в связи со стадией развития, на которой произошло раздвоение, и соответственно стадией закладки того или иного органа или признака. Зеркальность асимметричных признаков не может иногда не зависеть от стадии развития, на которой произошло раздвоение (Verschuer, 1929). К сожалению, этот вопрос до сих пор еще очень мало разработан.
Таким образом, по отношению к соединенным ОБ гипотеза Ньюмена кажется несколько более обоснованной, чем по отношению к свободным ОБ. Однако достаточно доказанной ее все же считать нельзя, и для полного выяснения правильности её или степени правильности необходимы дальнейшие исследования. Во всяком случае, в качестве рабочей гипотезы она была полезна и остается таковой и в настоящее время. Многие вопросы эта гипотеза до сих пор не может объяснить. Сам Ньюмен (Newman, 1940b) указал некоторые из них. Почему среди ОБ могут чаще встречаться признаки зеркальности, чем среди одиночек? Почему у ОБ одни признаки оказываются зеркальными, а другие нет — например при s. i. у одного из близнецов теменной завиток у него идет нормально? Почему у одних свободных ОБ с одинаковой степенью сходства зеркальность выступает резче, а у других слабее? Почему у одинаково соединенных близнецов может быть разное выражение зеркальности? Нет надобности увеличивать число таких вопросов; приведенных достаточно, чтобы показать, насколько вопрос о зеркальности у близнецов еще неясен, как много еще непонятного в нем.
Другого рода гипотезы были высказаны для объяснения зеркальности асимметричных признаков на основе генетики. В области наследственности асимметрии есть несколько интересных исследований, например Пшибрама (Phzibram, 1908) о наследственности разного цвета правого и левого глаза у ангорских кошек или Бойкотта с сотрудниками (Boycott et al., 1929) над улитками, имеющими спирально завитую раковину направо или налево. Объяснение, даваемое этим явлениям, сводилось на менделирование определенных генов. Дальберг (Dahlberg, 1926, 1929, 1943/44) по аналогии с этими явлениями высказал предположение о существовании неодинакового распределения генов при образовании ОБ, отчего зависит зеркальность асимметричных признаков, назвав свою гипотезу «теорией генотипической асимметрии». Эта гипотеза носит умозрительный характер и недостаточно подтверждается фактами. Несколько иначе, но в том же направлении Боутервек (Bouterwek, 1943) старался доказать, что асимметрия и зеркальность ОБ основаны на различии их генотипов. Этот тезис и его доказательства вызвали ряд возражений (Lotze, 1937, и др.), и действительно, попытки Боутервека не производят убедительного впечатления. Вопрос о значении генотипа для зеркальности у близнецов остается пока что нерешенным. Генотип, вероятно, играет известную роль и, может быть, неодинаковую у разных групп животных (Torgersen, 1950). Однако понять роль генотипа можно будет только тогда, когда глубже будет изучен физиологический механизм становления асимметрии зародыша, изменение этой асимметрии в зависимости от раздвоения зародыша и в связи с этим возможности возникновения зеркальности на разных ступенях развития эмбриона. Тогда будет также изжит односторонний характер гипотезы Ньюмена.
Теперь, после рассмотрения некоторых общих вопросов близнецовой проблемы, мы можем перейти к знакомству с многообразным проявлением близнечества в различных классах животного царства, начиная с низших форм и кончая человеком.
Глава восьмая БЛИЗНЕЦЫ У ЖИВОТНЫХ
Беспозвоночные
Мы начнем рассмотрение образования близнецов в этой обширной группе с представителей низшего типа многоклеточных животных — кишечнополостных. У гидры ОБ до последнего времени не были описаны. Совсем недавно установлены единичные случаи вылупления из эмбриотеки «двухголовых» эмбрионов, которых, очевидно, надо считать ОБ (рис. 64; Грузова, 1956).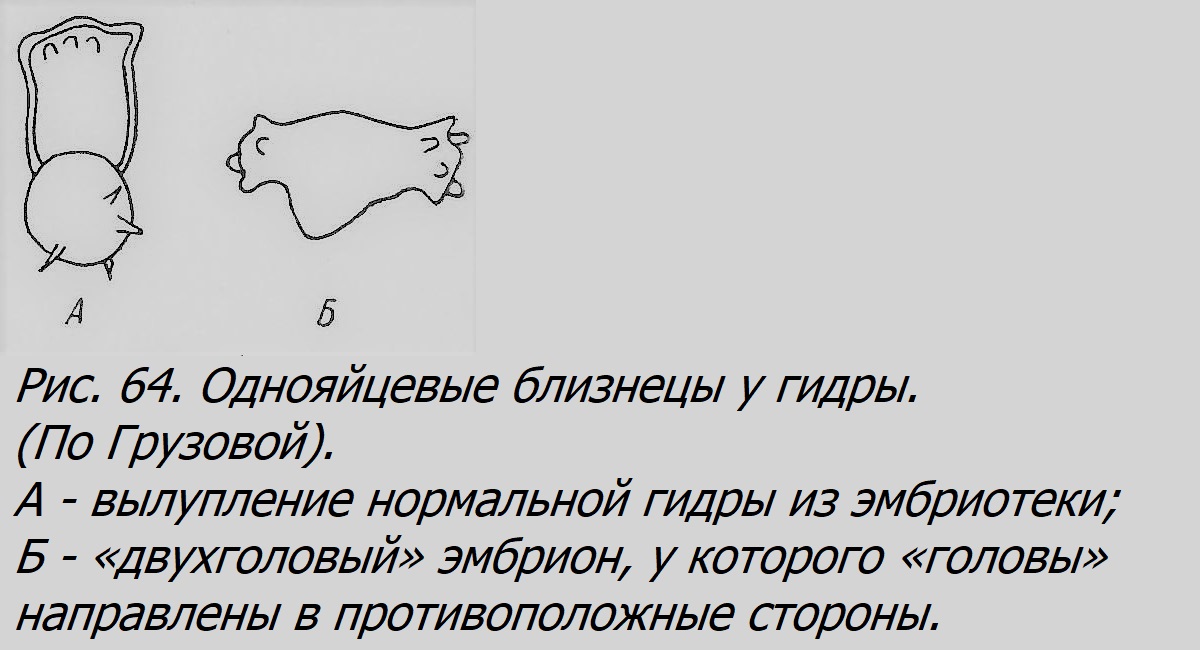 До известной степени аналогичное явление установлено на почках гидры, причем особи, возникшие из «близнецовых» почек, в свою очередь образовывал и такие почки приблизительно в два раза чаще, чем особи, возникшие из обыкновенных почек (Turner, 1956).Конечно, о почках-«близнецах» можно говорить только с известным насилием над, термином «близнецы». Вообще же вопрос о близнецах у гидр еще недостаточно выяснен и должен быть обстоятельно изучен. Но у некоторых других форм кишечнополостных ОБ получены экспериментально. Так, если начавшую дробиться зиготу (оплодотворенное яйцо) гидроида клитии (Clytia flavidula) разрезать тонкой острой иглой пополам на стадии первых двух клеток (бластомеров), т. е. полностью изолировать их друг от друга, то каждый из этих двух бластомеров будет продолжать дробиться, сначала как бы в контакте с другим, а потом, закругляясь, образует самостоятельный комплекс клеток. В процессе дальнейшего развития получается бластула[9], гаструла и, наконец, молодой полип, но только половинного против нормы размера (рис. 65).
До известной степени аналогичное явление установлено на почках гидры, причем особи, возникшие из «близнецовых» почек, в свою очередь образовывал и такие почки приблизительно в два раза чаще, чем особи, возникшие из обыкновенных почек (Turner, 1956).Конечно, о почках-«близнецах» можно говорить только с известным насилием над, термином «близнецы». Вообще же вопрос о близнецах у гидр еще недостаточно выяснен и должен быть обстоятельно изучен. Но у некоторых других форм кишечнополостных ОБ получены экспериментально. Так, если начавшую дробиться зиготу (оплодотворенное яйцо) гидроида клитии (Clytia flavidula) разрезать тонкой острой иглой пополам на стадии первых двух клеток (бластомеров), т. е. полностью изолировать их друг от друга, то каждый из этих двух бластомеров будет продолжать дробиться, сначала как бы в контакте с другим, а потом, закругляясь, образует самостоятельный комплекс клеток. В процессе дальнейшего развития получается бластула[9], гаструла и, наконец, молодой полип, но только половинного против нормы размера (рис. 65).
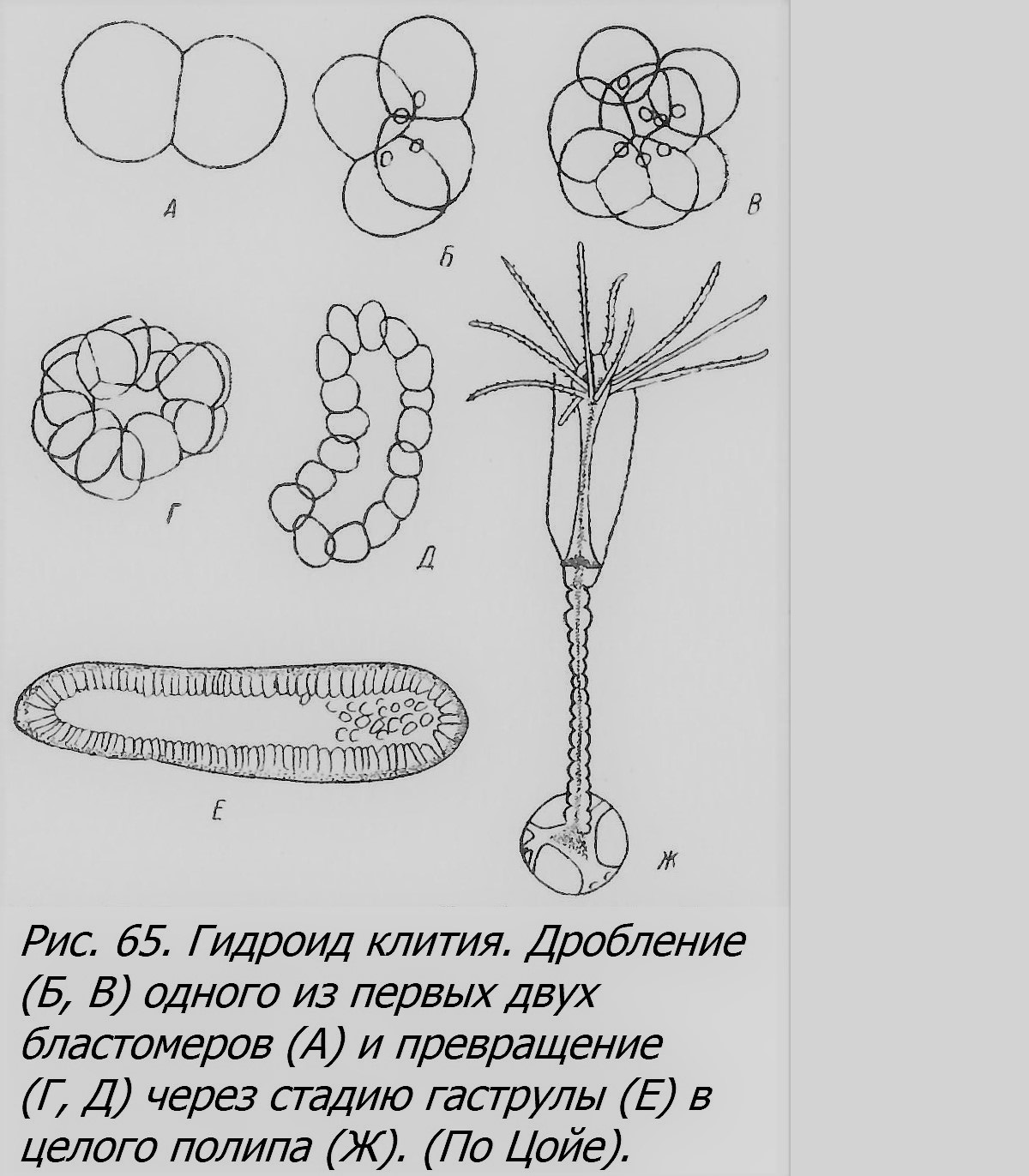 Если таким же способом изолировать первые четыре бластомера, то развитие пойдет аналогично, и получится четыре целых полипа, соответственно меньшего размера. И число щупалец у них вначале будет меньше обычного. Но уже при изоляции на стадии восьми бластомеров развитие каждого бластомера не идет и дальше гаструлы. Похожие результаты получены и на других гидроидах (Zoja, 1895), а также на высших кишечнополостных — сцифомедузах. Недавно колонии-близнецы получены у гидроидного полипа Hydractinia путем разделения первых двух бластомеров (Hauenschild, 1954).
Известно, что гидроиды и, в частности, пресноводные гидры в большой мере способны из небольших кусочков тела давать целое животное. Это явление регуляции можно до известной степени аналогизировать, с регуляцией отдельных бластомеров.
Яйца кишечнополостных нежны и легко разрушаются, с ними трудно ставить опыты и по ряду других причин.
Более удобным объектом для экспериментального получения близнецов являются иглокожие: морские ежи и морские звезды.Как и у гидроидов, близнецы у морских ежей получены путем изоляции бластомеров на самых ранних стадиях дробления яйца, стадиях двух и четырех бластомеров. На более поздних стадиях изолированные бластомеры уже не способны дать целый организм.
Изоляция достигается разными методами. Самый простой и грубый — это встряхивание сосуда, содержащего воду с дробящимися яйцами морского ежа (Driesch, 1892). Лёб (Loeb, 1894) помещал яйца морского ежа через 10 минут после оплодотворения в морскую воду, наполовину разведенную пресной водой. Яйца разбухали и лопались. Часть плазмы вытекала и закруглялась, не теряя связи с яйцом. После перенесения яиц в нормальную морскую воду развитие продолжалось; вытекшая плазма получала в процессе дробления ядро, и из нее тоже развивался зародыш, т. е. возникала пара близнецов, иногда тройня или четверня. Изящнее способ Гербста (Herbst, 1900): в искусственной морской воде, лишенной солей кальция, слой эктоплазмы растворяется, и бластомеры легко отделяются друг от друга, после чего их возвращают в нормальную морскую воду. Этот способ был развит и видоизменен Лёбом (Loeb, 1909), получавшим до 90% ОБ, частью симметричных (рис. 66),
Если таким же способом изолировать первые четыре бластомера, то развитие пойдет аналогично, и получится четыре целых полипа, соответственно меньшего размера. И число щупалец у них вначале будет меньше обычного. Но уже при изоляции на стадии восьми бластомеров развитие каждого бластомера не идет и дальше гаструлы. Похожие результаты получены и на других гидроидах (Zoja, 1895), а также на высших кишечнополостных — сцифомедузах. Недавно колонии-близнецы получены у гидроидного полипа Hydractinia путем разделения первых двух бластомеров (Hauenschild, 1954).
Известно, что гидроиды и, в частности, пресноводные гидры в большой мере способны из небольших кусочков тела давать целое животное. Это явление регуляции можно до известной степени аналогизировать, с регуляцией отдельных бластомеров.
Яйца кишечнополостных нежны и легко разрушаются, с ними трудно ставить опыты и по ряду других причин.
Более удобным объектом для экспериментального получения близнецов являются иглокожие: морские ежи и морские звезды.Как и у гидроидов, близнецы у морских ежей получены путем изоляции бластомеров на самых ранних стадиях дробления яйца, стадиях двух и четырех бластомеров. На более поздних стадиях изолированные бластомеры уже не способны дать целый организм.
Изоляция достигается разными методами. Самый простой и грубый — это встряхивание сосуда, содержащего воду с дробящимися яйцами морского ежа (Driesch, 1892). Лёб (Loeb, 1894) помещал яйца морского ежа через 10 минут после оплодотворения в морскую воду, наполовину разведенную пресной водой. Яйца разбухали и лопались. Часть плазмы вытекала и закруглялась, не теряя связи с яйцом. После перенесения яиц в нормальную морскую воду развитие продолжалось; вытекшая плазма получала в процессе дробления ядро, и из нее тоже развивался зародыш, т. е. возникала пара близнецов, иногда тройня или четверня. Изящнее способ Гербста (Herbst, 1900): в искусственной морской воде, лишенной солей кальция, слой эктоплазмы растворяется, и бластомеры легко отделяются друг от друга, после чего их возвращают в нормальную морскую воду. Этот способ был развит и видоизменен Лёбом (Loeb, 1909), получавшим до 90% ОБ, частью симметричных (рис. 66),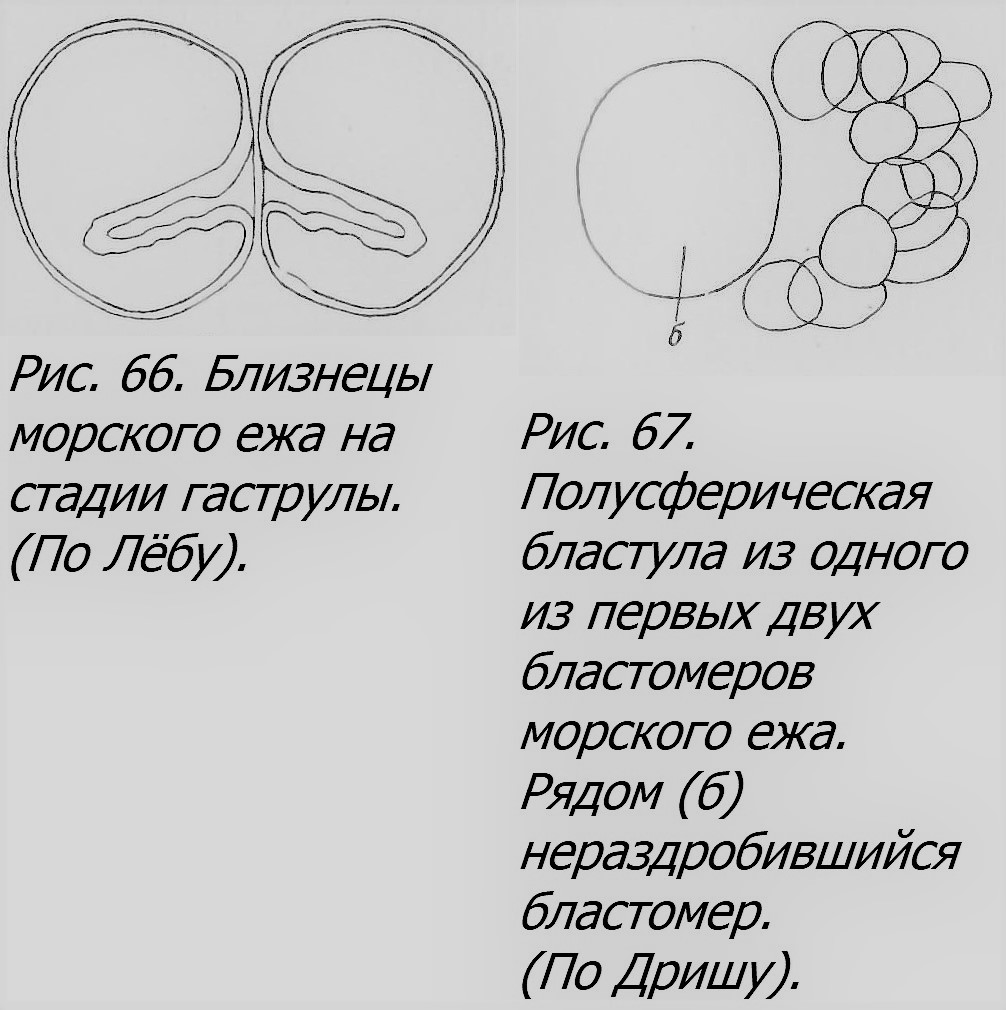 так как они возникали из первой пары бластомеров, физиологически изолированных. Существуют и другие способы (Harvey, 1940). Интересно отметить, что у некоторых видов морских ежен каждый из изолированных первых двух бластомеров, дробясь, образует полусферическую бластулу, «открытую», как при нормальном дроблении, когда рядом с ней другую полусферу образует парный с ней бластомер (рис. 67). Лишь при дальнейшем развитии бластула, замыкаясь, приобретает сферическую форму, а затем переходит в гаструлу и далее в личинку (плутеус), нормальной формы, но половинного размера (рис. 68).
так как они возникали из первой пары бластомеров, физиологически изолированных. Существуют и другие способы (Harvey, 1940). Интересно отметить, что у некоторых видов морских ежен каждый из изолированных первых двух бластомеров, дробясь, образует полусферическую бластулу, «открытую», как при нормальном дроблении, когда рядом с ней другую полусферу образует парный с ней бластомер (рис. 67). Лишь при дальнейшем развитии бластула, замыкаясь, приобретает сферическую форму, а затем переходит в гаструлу и далее в личинку (плутеус), нормальной формы, но половинного размера (рис. 68).
 Близнецы могут некоторое время развиваться, будучи слабосоединенными слизистой массой наружной поверхности их тел; но, начав свободно плавать, бластулы-двойни скоро разъединяются. Развитие близнецов у морских ежей может идти с разной скоростью, иногда с возникновениями разных аномалий. Как правило, по-видимому, развитие протекает тем медленнее, чем меньше размеры зародыша, в связи с тем, что он образовался из половины яйца (Horstadius, 1928 ). Аналогичное явление обнаружено у ланцетника (Morgan, 1927), тритона (Spemann u. Falkenberg, 1919) и у других обектов. Замедление развития наблюдается и у нормальных особей и, по-видимому, зависит преимущественно от различных внешних влияний на ход развития. Нет никаких данных для допущения, что между первыми двумя бластомерами было бы какое-либо существенное различие, что они, например, уже предназначены стать правой и левой половиной будущего зародыша. То же можно сказать и относительно первых четырех бластомеров — они, по-видимому, равноценны; это видно из того, что из них получаются целые зародыши-четверни.
В том случае, если из четырех бластомеров два обособятся, и два останутся вместе, получается тройня. Разумеется, зародыш из двух бластомеров окажется вдвое больше, чем каждый из двух других, возникших из одного бластомера.
В некоторых случаях два первых бластомера, сначала разошедшиеся, иногда снова соединяются. При этом могут наблюдаться разные степени слияния двух самостоятельно начавших развиваться зародышей. Получаются «сросшиеся» близнецы, иногда не отличимые, по-видимому, от целой особи, а в ряде случаев оказывающиеся с различными уродствами. Эти явления вторичного соединения зародышей у морских ежей еще мало изучены. Так же мало известны «свободные» близнецы этих животных, возникшие в природе.
Изолированные бластомеры на стадии восьми бластомеров у иглокожих уже не способны стать целым зародышем. Возникновение близнецов у морских ежей на более поздних стадиях развития, например гаструлы, еще недостаточно изучено.
Описаны «двойные яйца», образующиеся в тех случаях, когда второе редукционное тельце оказывается ненормально большого размера, иногда достигая размеров яйца. Из такого двойного яйца могут получиться близнецы (Lindahl, 1937). Но их нельзя называть однояйцевыми, как это делает Линдаль, так как, в сущности, здесь мы имеем дело уже с двумя разными яйцами, произошедшими из одного ооцита, аналогично тому, как образуются сперматозоиды, и, следовательно, близнецы, развивающиеся из такого «двойного яйца», разнояйцевые, оплодотворенные разными спермиями.
Подробно изучено образование близнецов у одного вида морских звезд (Patiria miniota). Ньюмен (Newman, 1921, 1923) различает три основных типа близнецов у этих животных: первый — «карликовые» личинки, получившиеся при физиологической, а далее и физической изоляции бластомеров, т. е. на ранней стадии дробления яйца; второй тип — двойни, тройни и т. д., возникшие на стадии гаструляции, и третий — «двухголовые» формы, образовавшиеся путем раздвоения переднего отдела первичной кишки. Все эти типы близнецов получаются, как думает Ньюмен, вследствие задержки развития, которая может быть вызвана разными причинами: партеногенезом, когда развитие идет вяло, медленно, оплодотворением чужой спермой (другого вида) и, наконец, неблагоприятными условиями развития, например теснотой, когда зародыши испытывают недостаток кислорода при избытке углекислого газа. Чаще других наблюдалось появление второго типа близнецов из нормально оплодотворенных яиц. Бластула, образующаяся из таких яиц, имеет перед гаструляцией явно выраженную полярность — у вегетативного полюса стенка бластулы заметно утолщена: в этом месте начинается впячивание стенки бластулы при гаструляции (рис. 69, А и Б).
Близнецы могут некоторое время развиваться, будучи слабосоединенными слизистой массой наружной поверхности их тел; но, начав свободно плавать, бластулы-двойни скоро разъединяются. Развитие близнецов у морских ежей может идти с разной скоростью, иногда с возникновениями разных аномалий. Как правило, по-видимому, развитие протекает тем медленнее, чем меньше размеры зародыша, в связи с тем, что он образовался из половины яйца (Horstadius, 1928 ). Аналогичное явление обнаружено у ланцетника (Morgan, 1927), тритона (Spemann u. Falkenberg, 1919) и у других обектов. Замедление развития наблюдается и у нормальных особей и, по-видимому, зависит преимущественно от различных внешних влияний на ход развития. Нет никаких данных для допущения, что между первыми двумя бластомерами было бы какое-либо существенное различие, что они, например, уже предназначены стать правой и левой половиной будущего зародыша. То же можно сказать и относительно первых четырех бластомеров — они, по-видимому, равноценны; это видно из того, что из них получаются целые зародыши-четверни.
В том случае, если из четырех бластомеров два обособятся, и два останутся вместе, получается тройня. Разумеется, зародыш из двух бластомеров окажется вдвое больше, чем каждый из двух других, возникших из одного бластомера.
В некоторых случаях два первых бластомера, сначала разошедшиеся, иногда снова соединяются. При этом могут наблюдаться разные степени слияния двух самостоятельно начавших развиваться зародышей. Получаются «сросшиеся» близнецы, иногда не отличимые, по-видимому, от целой особи, а в ряде случаев оказывающиеся с различными уродствами. Эти явления вторичного соединения зародышей у морских ежей еще мало изучены. Так же мало известны «свободные» близнецы этих животных, возникшие в природе.
Изолированные бластомеры на стадии восьми бластомеров у иглокожих уже не способны стать целым зародышем. Возникновение близнецов у морских ежей на более поздних стадиях развития, например гаструлы, еще недостаточно изучено.
Описаны «двойные яйца», образующиеся в тех случаях, когда второе редукционное тельце оказывается ненормально большого размера, иногда достигая размеров яйца. Из такого двойного яйца могут получиться близнецы (Lindahl, 1937). Но их нельзя называть однояйцевыми, как это делает Линдаль, так как, в сущности, здесь мы имеем дело уже с двумя разными яйцами, произошедшими из одного ооцита, аналогично тому, как образуются сперматозоиды, и, следовательно, близнецы, развивающиеся из такого «двойного яйца», разнояйцевые, оплодотворенные разными спермиями.
Подробно изучено образование близнецов у одного вида морских звезд (Patiria miniota). Ньюмен (Newman, 1921, 1923) различает три основных типа близнецов у этих животных: первый — «карликовые» личинки, получившиеся при физиологической, а далее и физической изоляции бластомеров, т. е. на ранней стадии дробления яйца; второй тип — двойни, тройни и т. д., возникшие на стадии гаструляции, и третий — «двухголовые» формы, образовавшиеся путем раздвоения переднего отдела первичной кишки. Все эти типы близнецов получаются, как думает Ньюмен, вследствие задержки развития, которая может быть вызвана разными причинами: партеногенезом, когда развитие идет вяло, медленно, оплодотворением чужой спермой (другого вида) и, наконец, неблагоприятными условиями развития, например теснотой, когда зародыши испытывают недостаток кислорода при избытке углекислого газа. Чаще других наблюдалось появление второго типа близнецов из нормально оплодотворенных яиц. Бластула, образующаяся из таких яиц, имеет перед гаструляцией явно выраженную полярность — у вегетативного полюса стенка бластулы заметно утолщена: в этом месте начинается впячивание стенки бластулы при гаструляции (рис. 69, А и Б).

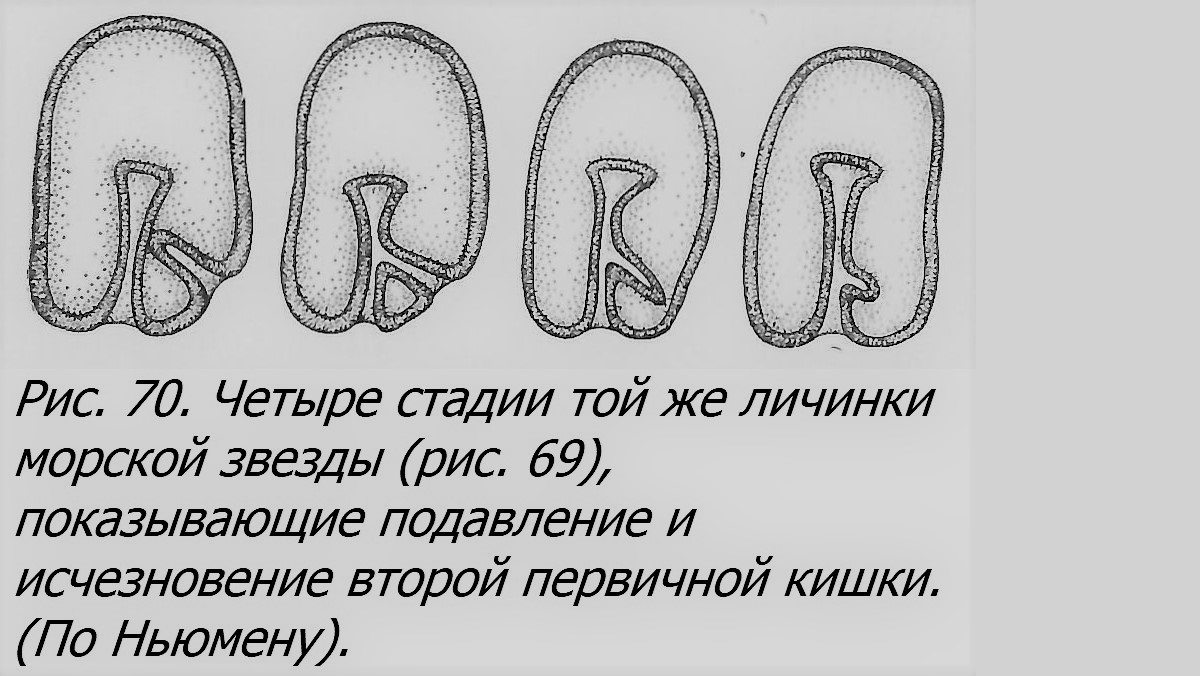
 Когда поляризация бластулы из-за неблагоприятных условий нарушена, то и гаструляция нарушается; впячивание первичной кишки идет одновременно в разных местах, иногда в двух противоположных направлениях (рис. 69, Е). Чаще всего образуется основная кишка, более мощная, и добавочная (рис. 70). В некоторых случая гаструляция носит симметричный характер, причем закладываются два как бы равноценных индивидуума. Основные оси их тела, поскольку эти оси определяются направлением роста первичной кишки, могут быть различно расположены в отношении друг друга, но симметрично в каждом случае, как бы зеркально (рис. 71).
В качестве вторичного явления возможно в дальнейшем соединение, в очень разных вариантах, двух первоначальных индивидуумов, срастание их. Так, например, первоначально две разные первичные кишки, растущие под острым углом друг к другу, сливаются и образуют в переднем конце одну общую кишку, а в заднем — две разные (рис. 71). Если же один из партнеров будет крупнее и сильнее другого, то последний оказывается постепенно подавленным первым и как бы поглощенным им (рис. 70). Обратив внимание лишь на одну из последних стадий, ее можно ошибочно принять за «почкование», а образование добавочной ветви из основной кишки. Это явление подавления одного близнеца другим у морской звезды Ньюмен сравнивает с похожими отношениями у позвоночных, когда на развитом организме (автозите) живет «вросший» в него близнец его (паразит), в разных случаях в различной степени недоразвитый и искаженный, своеобразный патологический придаток, обычно неотделимый.
Двойные уродства вроде описанных, однако, не всегда образуются путем такого соединения. В ряде случаев они, как допускает Ньюмен, возникают путем расщепления первоначально одного индивида — например так появляются особи раздвоенным передним концом, которых Ньюмен сравнивает с похожими явлениями у позвоночных (рис. 52).
Причина таких раздвоений у личинок морских звезд недостаточно изучена; ОБ встречаются чаще у отстающих в развитии особей, и, вероятно, это тоже является следствием временной физиологической изоляции соответственных двух участков организма, образовавших два самостоятельных пункта развития. Эти случаи, по Ньюмену, составляют третий тип образования близнецов у морских звезд.
Обращаясь к образованию близнецов у кольчатых червей, мы встречаемся у одного из видов их, Lumbriculus trapezoides[10], с замечательным явлением постоянного естественного образования однояйцевых близнецов, напоминающим явления такого рода у некоторых высших животных, например у армадилов (см. далее «Млекопитающие»), однако с существенным различием в ходе процесса раздвоения эмбриона. Названный вид земляных червей откладывает в кокон несколько яиц, из которых чаще всего развивается одно. Яйцо сначала делится на два крупных бластомера и далее на шесть мелких, располагающихся между крупными (рис. 72).
Когда поляризация бластулы из-за неблагоприятных условий нарушена, то и гаструляция нарушается; впячивание первичной кишки идет одновременно в разных местах, иногда в двух противоположных направлениях (рис. 69, Е). Чаще всего образуется основная кишка, более мощная, и добавочная (рис. 70). В некоторых случая гаструляция носит симметричный характер, причем закладываются два как бы равноценных индивидуума. Основные оси их тела, поскольку эти оси определяются направлением роста первичной кишки, могут быть различно расположены в отношении друг друга, но симметрично в каждом случае, как бы зеркально (рис. 71).
В качестве вторичного явления возможно в дальнейшем соединение, в очень разных вариантах, двух первоначальных индивидуумов, срастание их. Так, например, первоначально две разные первичные кишки, растущие под острым углом друг к другу, сливаются и образуют в переднем конце одну общую кишку, а в заднем — две разные (рис. 71). Если же один из партнеров будет крупнее и сильнее другого, то последний оказывается постепенно подавленным первым и как бы поглощенным им (рис. 70). Обратив внимание лишь на одну из последних стадий, ее можно ошибочно принять за «почкование», а образование добавочной ветви из основной кишки. Это явление подавления одного близнеца другим у морской звезды Ньюмен сравнивает с похожими отношениями у позвоночных, когда на развитом организме (автозите) живет «вросший» в него близнец его (паразит), в разных случаях в различной степени недоразвитый и искаженный, своеобразный патологический придаток, обычно неотделимый.
Двойные уродства вроде описанных, однако, не всегда образуются путем такого соединения. В ряде случаев они, как допускает Ньюмен, возникают путем расщепления первоначально одного индивида — например так появляются особи раздвоенным передним концом, которых Ньюмен сравнивает с похожими явлениями у позвоночных (рис. 52).
Причина таких раздвоений у личинок морских звезд недостаточно изучена; ОБ встречаются чаще у отстающих в развитии особей, и, вероятно, это тоже является следствием временной физиологической изоляции соответственных двух участков организма, образовавших два самостоятельных пункта развития. Эти случаи, по Ньюмену, составляют третий тип образования близнецов у морских звезд.
Обращаясь к образованию близнецов у кольчатых червей, мы встречаемся у одного из видов их, Lumbriculus trapezoides[10], с замечательным явлением постоянного естественного образования однояйцевых близнецов, напоминающим явления такого рода у некоторых высших животных, например у армадилов (см. далее «Млекопитающие»), однако с существенным различием в ходе процесса раздвоения эмбриона. Названный вид земляных червей откладывает в кокон несколько яиц, из которых чаще всего развивается одно. Яйцо сначала делится на два крупных бластомера и далее на шесть мелких, располагающихся между крупными (рис. 72).
 Дальнейшее дробление, также неравномерное, приводит к образованию бластулы. Образование гаструлы начинается инвагинацией двух крупных клеток, с последующей дифференцировкой правой и левой стороны зародыша. Яйцо удлиняется, появляется поперечная борозда, разделяющая яйцо пополам. Так возникают близнецы, которые могут оставаться в той или иной степени и форме соединенными или же разойтись вовсе, став самостоятельными организмами, что происходит обычно при первых движениях близнецов (Кleinenberg, 1879).
Двойные уродства возникают, надо думать, вследствие толщины связующей близнецов клеточной массы, мешающей им разойтись. Иногда партнеры развиваются не одинаково — один отстает от другого, оказывается угнетенным и может достигать положения «паразита» на опередившем его, более мощном близнеце.
Двойные уродства описаны и у других видов земляных червей (например, Allobophora subrubicunda). Партнеры в таких случаях чаще соединены средней частью тела, а передний или задний конец, а иногда и тот и другой, в разной степени раздвоены (рис. 73) (Korschelt, 1904).
Дальнейшее дробление, также неравномерное, приводит к образованию бластулы. Образование гаструлы начинается инвагинацией двух крупных клеток, с последующей дифференцировкой правой и левой стороны зародыша. Яйцо удлиняется, появляется поперечная борозда, разделяющая яйцо пополам. Так возникают близнецы, которые могут оставаться в той или иной степени и форме соединенными или же разойтись вовсе, став самостоятельными организмами, что происходит обычно при первых движениях близнецов (Кleinenberg, 1879).
Двойные уродства возникают, надо думать, вследствие толщины связующей близнецов клеточной массы, мешающей им разойтись. Иногда партнеры развиваются не одинаково — один отстает от другого, оказывается угнетенным и может достигать положения «паразита» на опередившем его, более мощном близнеце.
Двойные уродства описаны и у других видов земляных червей (например, Allobophora subrubicunda). Партнеры в таких случаях чаще соединены средней частью тела, а передний или задний конец, а иногда и тот и другой, в разной степени раздвоены (рис. 73) (Korschelt, 1904).
 Как известно, такие же уродства можно получать искусственно, путем надреза переднего или заднего конца такого червя; это свидетельствует о том, что изоляция ведет к обособлению соответствующей части в процессе регенерации. Вероятно, в естественных условиях различные факторы среды могут вызывать физиологическую изоляцию соответствующих отделов зародыша, что имеет следствием его раздвоение. Но этот вопрос на червях еще мало изучен экспериментально.
Интересно отметить, что существуют другие виды земляных червей, родственные данным видам, которые очень редко образуют близнецов (Vejdovsky, 1882—1892).
Среди мшанок, этих своеобразных водных колониальных животных, есть виды, обладающие способностью к регулярному размножению путем образования ОБ, возникающих на ранней стадии зародыша, до бластулы; происходит распад его на значительное количество частей, из которых каждая становится самостоятельным эмбрионом (Caullery, 1945, и др.).
Как известно, такие же уродства можно получать искусственно, путем надреза переднего или заднего конца такого червя; это свидетельствует о том, что изоляция ведет к обособлению соответствующей части в процессе регенерации. Вероятно, в естественных условиях различные факторы среды могут вызывать физиологическую изоляцию соответствующих отделов зародыша, что имеет следствием его раздвоение. Но этот вопрос на червях еще мало изучен экспериментально.
Интересно отметить, что существуют другие виды земляных червей, родственные данным видам, которые очень редко образуют близнецов (Vejdovsky, 1882—1892).
Среди мшанок, этих своеобразных водных колониальных животных, есть виды, обладающие способностью к регулярному размножению путем образования ОБ, возникающих на ранней стадии зародыша, до бластулы; происходит распад его на значительное количество частей, из которых каждая становится самостоятельным эмбрионом (Caullery, 1945, и др.).
 Переходим к членистоногим. У скорпионов (Brauer, 1917), представителей класса паукообразных, описаны как свободные ОБ (рис. 74, А, Б),так и некоторые основные типы соединенных близнецов: переднее раздвоение (рис. 74, В), заднее раздвоение (рис. 74, Д) и «янусообразное» уродство, известное у низших позвоночных — тритонов (см. далее «Амфибии») и у человека (см. главу шестую). Эти двойные уроды, найденные в природе, вполне аналогичны полученным экспериментально у тритонов, и, вероятно, возникновение их должно объясняться такими же процессами, которые наблюдались у амфибий. По-видимому, такие «янусы», двуликие уродства, возникают на стадии гаструляции, когда два «потока» тканей, двигаясь навстречу друг другу, сталкиваются и расходятся в стороны под прямым углом. Благодаря этому передние части компонентов оказываются слившимися наполовину из одного потока, а наполовину из другого. Возникновение двух очагов развития в одном яйце, из которых и происходит такой встречный рост, подтверждается Брауером описанием некоторых яиц, где такая картина видна (рис. 74, Е), напоминая описанную Ветцелем на яйце змеи (см. далее «Рептилии»).
Хорошо изучены близнецы у некоторых представителей паразитических насекомых из отряда перепончатокрылых. Эти животные откладывают свои яйца на тело других насекомых, на их яйца и т. д. Вероятно, с паразитизмом сопряжено у них и развитие способности яйца превращаться не в одного зародыша, а в несколько; у некоторых видов из одного яйца получается свыше тысячи зародышей, иначе говоря, ОБ; например, у Litomastrix truncatellus получается около 1480 зародышей из одного яйца (Silvestri, 1906). Близнецы получаются путем распада яйца на несколько несимметричных обособленных участков, из которых каждый развивается дальше, образуя самостоятельный зародыш. Этим процесс образования ОБ у данных насекомых отличается от такового у позвоночных, где эмбрион чаще всего делится на пару симметричных особей. В качестве примера полиэмбрионии у перепончатокрылых может служить Platygaster vernalis, откладывающий свои яйца на яйца гессенской мухи. Ядро оплодотворенного яйца платигастера повторно делится на 16 частей (рис. 75), каждое из ядер окружается слоем цитоплазмы, и далее но числу ядер образуются отдельные клетки. Из каждой клетки путем ее дробления может получиться один или два эмбриона; обычно не все эмбрионы развиваются до конца, часть их гибнет. В среднем из одного яйца получается 8 зародышей-близнецов (Patterson, 1927).
Переходим к членистоногим. У скорпионов (Brauer, 1917), представителей класса паукообразных, описаны как свободные ОБ (рис. 74, А, Б),так и некоторые основные типы соединенных близнецов: переднее раздвоение (рис. 74, В), заднее раздвоение (рис. 74, Д) и «янусообразное» уродство, известное у низших позвоночных — тритонов (см. далее «Амфибии») и у человека (см. главу шестую). Эти двойные уроды, найденные в природе, вполне аналогичны полученным экспериментально у тритонов, и, вероятно, возникновение их должно объясняться такими же процессами, которые наблюдались у амфибий. По-видимому, такие «янусы», двуликие уродства, возникают на стадии гаструляции, когда два «потока» тканей, двигаясь навстречу друг другу, сталкиваются и расходятся в стороны под прямым углом. Благодаря этому передние части компонентов оказываются слившимися наполовину из одного потока, а наполовину из другого. Возникновение двух очагов развития в одном яйце, из которых и происходит такой встречный рост, подтверждается Брауером описанием некоторых яиц, где такая картина видна (рис. 74, Е), напоминая описанную Ветцелем на яйце змеи (см. далее «Рептилии»).
Хорошо изучены близнецы у некоторых представителей паразитических насекомых из отряда перепончатокрылых. Эти животные откладывают свои яйца на тело других насекомых, на их яйца и т. д. Вероятно, с паразитизмом сопряжено у них и развитие способности яйца превращаться не в одного зародыша, а в несколько; у некоторых видов из одного яйца получается свыше тысячи зародышей, иначе говоря, ОБ; например, у Litomastrix truncatellus получается около 1480 зародышей из одного яйца (Silvestri, 1906). Близнецы получаются путем распада яйца на несколько несимметричных обособленных участков, из которых каждый развивается дальше, образуя самостоятельный зародыш. Этим процесс образования ОБ у данных насекомых отличается от такового у позвоночных, где эмбрион чаще всего делится на пару симметричных особей. В качестве примера полиэмбрионии у перепончатокрылых может служить Platygaster vernalis, откладывающий свои яйца на яйца гессенской мухи. Ядро оплодотворенного яйца платигастера повторно делится на 16 частей (рис. 75), каждое из ядер окружается слоем цитоплазмы, и далее но числу ядер образуются отдельные клетки. Из каждой клетки путем ее дробления может получиться один или два эмбриона; обычно не все эмбрионы развиваются до конца, часть их гибнет. В среднем из одного яйца получается 8 зародышей-близнецов (Patterson, 1927).
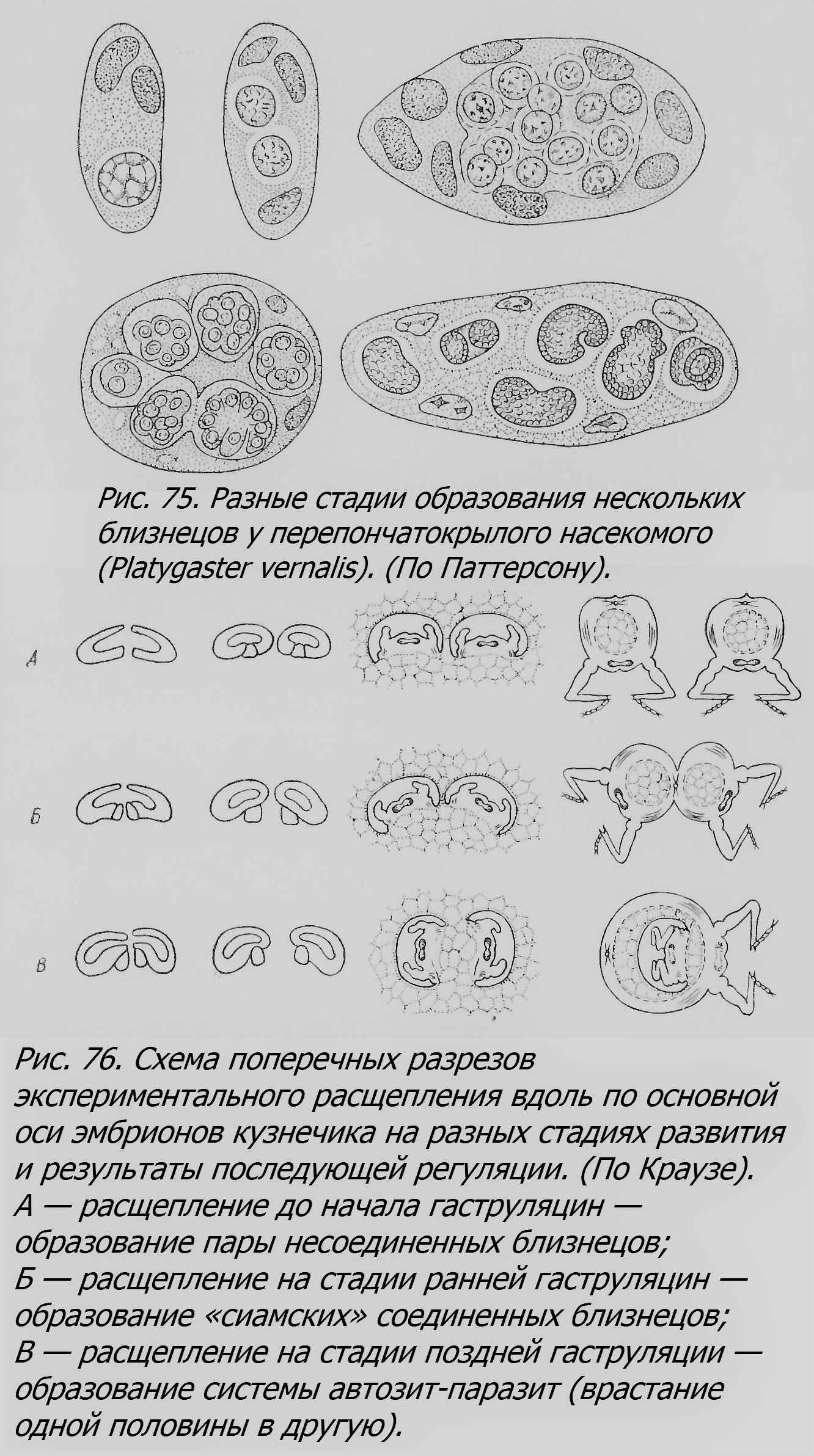 ОБ у насекомых были получены также экспериментальными методами. Например, тонкой микрохирургической техникой можно делать надрез в разных участках зародыша на различных стадиях развития — до гаструляции, во время нее и после (Krause, 1953). В разной степени изолированные участки эмбриона одного вида кузнечика в соответствии с условиями опыта дают многообразную картину удвоения, от вполне самостоятельных, свободных близнецов до различных степеней и форм двойных уродств (рис. 76).
Различные типы соединенных близнецов у насекомых, полученных в этих опытах, оказываются в ряде случаев аналогичными двойным уродствам у позвоночных, например симметричные «сиамские» близнецы или асимметричные (автозит-паразит; рис. 77). Способом надреза и вызванной им изоляции соответственных частей систематически изучались детерминированность и способность к перестройке различных участков зародыша насекомого на разных стадиях развития, что имеет методическое и теоретическое значение, на котором мы здесь останавливаться не можем.
ОБ у насекомых были получены также экспериментальными методами. Например, тонкой микрохирургической техникой можно делать надрез в разных участках зародыша на различных стадиях развития — до гаструляции, во время нее и после (Krause, 1953). В разной степени изолированные участки эмбриона одного вида кузнечика в соответствии с условиями опыта дают многообразную картину удвоения, от вполне самостоятельных, свободных близнецов до различных степеней и форм двойных уродств (рис. 76).
Различные типы соединенных близнецов у насекомых, полученных в этих опытах, оказываются в ряде случаев аналогичными двойным уродствам у позвоночных, например симметричные «сиамские» близнецы или асимметричные (автозит-паразит; рис. 77). Способом надреза и вызванной им изоляции соответственных частей систематически изучались детерминированность и способность к перестройке различных участков зародыша насекомого на разных стадиях развития, что имеет методическое и теоретическое значение, на котором мы здесь останавливаться не можем.
 Другой метод получения близнецов у насекомых построен на действии ядовитых веществ, подавляющих развитие. Так, например, на развивающееся яйцо одного жука (Bruchus quadrimaculatus) действуют раствором цианистого калия из расчета, согласно теории Чайлда, что яд прежде всего повлияет угнетающе на физиологически самую активную часть зародыша, каковой оказывается передняя часть будущей (презумитивной) брюшной стороны. По возвращении в нормальные условия развитие продолжается, но при этом возникают физиологически изолированные новые очаги развития, обычно два симметрично по обе стороны первоначального, уничтоженного действием яда. В результате этого зародыш раздваивается продольно (рис. 78). В зависимости от стадии развития и длительности воздействия цианистого калия можно вызвать разные степени раздвоения эмбриона, вплоть до образования вполне раздельных близнецов (Brauer, 1938).
Таким образом, у насекомых, как и у многих других беспозвоночных разных типов и классов, вследствие физической или физиологической изоляции участков яйца или зародыша на ранних стадиях развития, когда возможна еще регуляция, могут при известных условиях, естественных или экспериментальных, возникать ОБ.
Другой метод получения близнецов у насекомых построен на действии ядовитых веществ, подавляющих развитие. Так, например, на развивающееся яйцо одного жука (Bruchus quadrimaculatus) действуют раствором цианистого калия из расчета, согласно теории Чайлда, что яд прежде всего повлияет угнетающе на физиологически самую активную часть зародыша, каковой оказывается передняя часть будущей (презумитивной) брюшной стороны. По возвращении в нормальные условия развитие продолжается, но при этом возникают физиологически изолированные новые очаги развития, обычно два симметрично по обе стороны первоначального, уничтоженного действием яда. В результате этого зародыш раздваивается продольно (рис. 78). В зависимости от стадии развития и длительности воздействия цианистого калия можно вызвать разные степени раздвоения эмбриона, вплоть до образования вполне раздельных близнецов (Brauer, 1938).
Таким образом, у насекомых, как и у многих других беспозвоночных разных типов и классов, вследствие физической или физиологической изоляции участков яйца или зародыша на ранних стадиях развития, когда возможна еще регуляция, могут при известных условиях, естественных или экспериментальных, возникать ОБ.
Позвоночные
Низшие позвоночные
Прежде чем обратиться к рассмотрению близнецов у высших позвоночных, скажем несколько слов об экспериментальном получении ОБ у представителя низших, бесчерепных позвоночных, к которым относят ланцетника (Amphioxus). Его яйца претерпевают полное дробление, как и яйца морских ежей, почему у них можно так же легко получить изоляцию отдельных бластомеров на самых ранних стадиях дробления; это достигается тряской сосуда с водой, где находятся дробящиеся яйца ланцетника. Из каждого из первых двух бластомеров получается нормальный зародыш, т. е. ОБ. При неполном расхождении бластомеров возникают различные формы соединенных близнецов, обособленных друг от друга или с переднего конца (duplicitas anterior), или с заднего (d. posterior). Иногда получаются причудливые двойные уродства, например соединенные правыми боками, т. е. так, что головной конец одного расположен параллельно хвостовому другого. Такое соединение компонентов происходит из-за того, что вследствие тряски бластомеры могут не разойтись, а только сместиться по отношению друг к другу (Conklin, 1933).
У круглоротых, относящихся к бесчелюстным позвоночным, у которых дробление яйца также полное, можно получить ОБ и двойные уродства, если временно задержать дробление яиц, поместив их в гипертонический раствор сахара или морской соли. Этим приемом, по-видимому, достигается физиологическая изоляция бластомеров. Механическая изоляция первых двух бластомеров миноги тоже может давать ОБ (Caullery, 1945). Если после нормальной кладки яиц извлечь из животного задержавшиеся яйца, очевидно уже перезрелые, и искусственно оплодотворить их, то наблюдается раздвоение зародышей на стадии морулы или гаструлы, иногда до 40%. Вероятно, это объясняется возникновением физиологически изолированных очагов развития, как и у перезрелых яиц лягушки (см. дальше «Амфибии»).
Рыбы
Строение их яйца, как известно, отличается относительно большим запасом желтка, в связи с чем дробление у костистых рыб неполное, начинающееся с появления небольшого диска у анимального[11] полюса. Разрастаясь, этот диск, называемый бластодиском, постепенно охватывает недробящуюся массу желтка. На краю бластодиска уже рано возникает краевой валик, где клетки интенсивно размножаются. На краевом валике, пока диск еще невелик, возникает утолщение, краевой узелок; им определяется будущий задний конец, диска. Отсюда начинается закладка основной оси зародыша рыбы, причем становящаяся голова обращена к анимальному полюсу. У рыб, как и у беспозвоночных, близнецы возникают до и во время гаструляции. По окончании нее ОБ уже не образуются. У низших рыб, селяхий, описаны двойные уродства. У ската найдено яйцо с двумя зародышевыми дисками (Caullery, 1945). Таких фактов немного, и близнецы у этой группы рыб мало изучены.
 Гораздо лучше исследованы близнецы у высших рыб, костистых, как найденные в природе, где они в общем довольно редко встречаются, так и полученные в эксперименте. ОБ у костистых рыб могут возникать, по-видимому, по крайней мере двумя путями: 1) путем образования добавочных краевых узелков, т. е. пунктов, откуда начинается гаструляция, и 2) путем расщепления надвое по продольной оси первоначально одного эмбриона. Первый путь, по-видимому, имеет несколько вариантов. Могут оказаться физиологически изолированными группы бластомеров (рис. 79), которые, возможно, дадут начало разным зародышевым дискам. Могут возникнуть разные точки гаструляция, в разной мере удаленные друг от друга на бластодиске (рис. 80). Если они закладываются на противоположных краях диска и растут навстречу друг другу, то получатся более или менее нормальные зародыши, соединенные только общим желточным мешком или тканями тела в области брюха (рис. 81). Не всегда партнеры оказываются равновеликими и равноценными. Наблюдаются различные ступени перехода к комбинациям, когда один из близнецов оказывается развитым относительно нормально, а другой — отставшим в развитии, с дефектами в формировании различных частей тела (головы, глаз, туловища, конечностей и т. д.), словом, здесь создаются отношения типа автозит-паразит (рис. 81, Б — Г). Близнецы рыб, соединенные в брюшной области, морфологически аналогичны близнецам млекопитающих и человека, которые называются торакопаги, и близким к ним формам.
Гораздо лучше исследованы близнецы у высших рыб, костистых, как найденные в природе, где они в общем довольно редко встречаются, так и полученные в эксперименте. ОБ у костистых рыб могут возникать, по-видимому, по крайней мере двумя путями: 1) путем образования добавочных краевых узелков, т. е. пунктов, откуда начинается гаструляция, и 2) путем расщепления надвое по продольной оси первоначально одного эмбриона. Первый путь, по-видимому, имеет несколько вариантов. Могут оказаться физиологически изолированными группы бластомеров (рис. 79), которые, возможно, дадут начало разным зародышевым дискам. Могут возникнуть разные точки гаструляция, в разной мере удаленные друг от друга на бластодиске (рис. 80). Если они закладываются на противоположных краях диска и растут навстречу друг другу, то получатся более или менее нормальные зародыши, соединенные только общим желточным мешком или тканями тела в области брюха (рис. 81). Не всегда партнеры оказываются равновеликими и равноценными. Наблюдаются различные ступени перехода к комбинациям, когда один из близнецов оказывается развитым относительно нормально, а другой — отставшим в развитии, с дефектами в формировании различных частей тела (головы, глаз, туловища, конечностей и т. д.), словом, здесь создаются отношения типа автозит-паразит (рис. 81, Б — Г). Близнецы рыб, соединенные в брюшной области, морфологически аналогичны близнецам млекопитающих и человека, которые называются торакопаги, и близким к ним формам.

 В случае, если закладка двух и более близнецов произойдет на диске по соседству друг от друга, то в процессе роста и развития зародыши могут сблизиться и в той или иной мере срастись, образуя как разные формы равновеликих соединенных близнецов — симметричных (рис. 82), так и неодинаково развитых, где тоже может выступить отношение автозит-паразит в разной степени контраста. В отличие от первого типа партнеры будут здесь соединены боками, чаще имея две разные головы, чем два хвоста; например, до 90% всех таких двойных уродств форели имеют две головы (Lynn, 1938). Это тип, уже нам встречавшийся, называемый duplicitas anterior, т. е. переднее раздвоение, в отличие от d. posterior — заднего раздвоения. Переднее раздвоение может быть различной степени: от едва заметного снаружи раздвоения головы при полной норме всего тела (рис. 83) до выраженного в той или иной мере раздвоения не только головы, по и переднего участка тела (рис. 84). Интересно, что в таких случаях у одного из партнеров, чаще правого, нередко внутренние органы лежат в порядке, обратном норме, зеркально отражая обычное расположение внутренностей, т. е. имеется situs inversus.
В случае, если закладка двух и более близнецов произойдет на диске по соседству друг от друга, то в процессе роста и развития зародыши могут сблизиться и в той или иной мере срастись, образуя как разные формы равновеликих соединенных близнецов — симметричных (рис. 82), так и неодинаково развитых, где тоже может выступить отношение автозит-паразит в разной степени контраста. В отличие от первого типа партнеры будут здесь соединены боками, чаще имея две разные головы, чем два хвоста; например, до 90% всех таких двойных уродств форели имеют две головы (Lynn, 1938). Это тип, уже нам встречавшийся, называемый duplicitas anterior, т. е. переднее раздвоение, в отличие от d. posterior — заднего раздвоения. Переднее раздвоение может быть различной степени: от едва заметного снаружи раздвоения головы при полной норме всего тела (рис. 83) до выраженного в той или иной мере раздвоения не только головы, по и переднего участка тела (рис. 84). Интересно, что в таких случаях у одного из партнеров, чаще правого, нередко внутренние органы лежат в порядке, обратном норме, зеркально отражая обычное расположение внутренностей, т. е. имеется situs inversus.
 Только что рассмотренный тип соединенных близнецов мы считали, вслед за Стокардом (Stockard, 1921) и другими, результатом вторичного слияния двух первоначально разных зародышей, возникших из одного яйца. Однако такие объекты некоторые авторы, например Ньюмен (Newmen, 1923), принимают за результат продольного расщепления первоначально одного эмбриона, относя их ко второму типу образования близнецов у рыб, о котором речь была выше. По аналогии с другими высшими позвоночными этот тип вероятен и у рыб, тем более, что есть факты, которые, по-видимому, прямо говорят в пользу его существования. Так, например, Бэр (1845а) описал два случая соединенных близнецов окуня, которые он решительно считал за продукты расщепления одного эмбриона, а не слияния двух. Один из этих двойных уродов был менее раздвоен, чем другой (рис. 15). Оба были вполне прозрачны и не имели никаких следов срастания. Бэр сравнивал их с аналогичной стадией раздвоения цыпленка, описанной им (Бэр, 1827). Конечно, в отношении некоторых конкретных объектов может быть окажется не легко решить, каким путем они возникли — первым или вторым. Но во всяком случае, оба варианта являются ОБ.
Близнецы у рыб были получены также экспериментально. Стокард (Stockard, 1921) действовал на ранние стадии развития яиц двух видов рыб — фундулуса и форели — факторами, задерживающими развитие: низкой температурой (до 6—8° С) и уменьшением содержания кислорода в воде, необходимого для интенсивного обмена дробящихся яиц. У обоих видов рыб временная задержка развития вызывала возникновение большего, чем обычно встречается, числа близнецов разного рода. Так, например, у фундулуса при развитии в нормальных условиях одна двойня встречается приблизительно на тысячу яиц, а в эксперименте с пониженной температурой в некоторых случаях число близнецов достигало 9.3% всех яиц. Помимо образования близнецов холод и удушье вызывают гибель значительной части яиц. Также возрастает число различных аномалий развития и уродств. Интересно, что яйца одной кладки одной и той же особи имеют, по-видимому, разную степень сопротивляемости по отношению к вредящим факторам. Одни яйца гибнут, другие дают двойни или уродства, а третьи развиваются нормально. Сущность этих различий еще недостаточно выяснена. Есть также видовые различия реакции на вредные факторы. Форель, например, дает заметно больший процент близнецов, чем фундулус. По-видимому, в яйцах форели легче возникает физиологическая изоляция отдельных участков под влиянием вредящих воздействий, чем в яйцах фундулуса, согласно представлению Стокарда. В зависимости от того, на какой стадии эмбриогенеза действовал вредящий фактор, происходит остановка развития тех органов, которые наиболее интенсивно развиваются на этой стадии, а в связи с этим — соответствующие дефекты развития этих органов (мозга, глаз, конечностей и т. д.). Вредящее, искажающее развитие влияние оказывает также на своего партнера более развитой компонент, автозит на паразита, который в свою очередь тоже не может быть безразличным для автозита.
Другая серия опытов с целью получения близнецов была предпринята на яйцах фундулуса, подвергавшихся минутному облучению ультрафиолетовыми лучами (Hinrichs a. Genther, 1931). Яйца облучались, начиная с 5 минут после оплодотворения. Наибольшее число близнецов получалось под влиянием облучения на 25—30-й минуте и достигало 8.9%. И в этих опытах образование близнецов сопровождалось множественной гибелью яиц и разнообразными уродствами. Облучение за первые полчаса после оплодотворения яиц приходилось на такой период их жизни, когда дробление еще не начиналось или только что начиналось и никакой видимой морфологической дифференцировки еще не было. Большинство близнецов возникало, несомненно,из одного бластодиска, разделившегося, как думают авторы, еще задолго до гаструляции. Образование двух бластодисков на одном яйце с достоверностью не установлено. Раннее облучение способствует раздвоению переднего конца эмбриона, более позднее — заднего, поскольку развитие начинается с головного конца. Интересны некоторые числовые отношения полученных близнецов при облучении не позже, чем через 20 минут после оплодотворения; например, из 106 близнецов было получено: 5 пар с нормальными головами, 21 пара с одной нормальной и одной ненормальной головой, 9 пар, состоящих из слабо дифференцированной массы, одна пара имела частично раздвоенное сердце и 70 пар имели признаки слияния по основной оси (удвоенные ряды сомитов[12] и т. п., — рис. 85). Большинство, таким образом, состояло из соединенных близнецов, получившихся, вероятно, путем слияния двух самостоятельно образовавшихся зачатков, расположенных сравнительно близко друг от друга на общем бластодиске. Гаструляция на противоположных сторонах диска, т. е. под углом 180°, встречается сравнительно редко (рис. 85, Л). Микроскопическое изучение близнецов фундулуса, возникших под влиянием облучения как свободных, так и соединенных, обнаружило зеркальное расположение органов (Hinrichs, 1938). Рисунки, к сожалению, в этой работе не даны, и описание очень кратко; процент s. i. не указан.
Только что рассмотренный тип соединенных близнецов мы считали, вслед за Стокардом (Stockard, 1921) и другими, результатом вторичного слияния двух первоначально разных зародышей, возникших из одного яйца. Однако такие объекты некоторые авторы, например Ньюмен (Newmen, 1923), принимают за результат продольного расщепления первоначально одного эмбриона, относя их ко второму типу образования близнецов у рыб, о котором речь была выше. По аналогии с другими высшими позвоночными этот тип вероятен и у рыб, тем более, что есть факты, которые, по-видимому, прямо говорят в пользу его существования. Так, например, Бэр (1845а) описал два случая соединенных близнецов окуня, которые он решительно считал за продукты расщепления одного эмбриона, а не слияния двух. Один из этих двойных уродов был менее раздвоен, чем другой (рис. 15). Оба были вполне прозрачны и не имели никаких следов срастания. Бэр сравнивал их с аналогичной стадией раздвоения цыпленка, описанной им (Бэр, 1827). Конечно, в отношении некоторых конкретных объектов может быть окажется не легко решить, каким путем они возникли — первым или вторым. Но во всяком случае, оба варианта являются ОБ.
Близнецы у рыб были получены также экспериментально. Стокард (Stockard, 1921) действовал на ранние стадии развития яиц двух видов рыб — фундулуса и форели — факторами, задерживающими развитие: низкой температурой (до 6—8° С) и уменьшением содержания кислорода в воде, необходимого для интенсивного обмена дробящихся яиц. У обоих видов рыб временная задержка развития вызывала возникновение большего, чем обычно встречается, числа близнецов разного рода. Так, например, у фундулуса при развитии в нормальных условиях одна двойня встречается приблизительно на тысячу яиц, а в эксперименте с пониженной температурой в некоторых случаях число близнецов достигало 9.3% всех яиц. Помимо образования близнецов холод и удушье вызывают гибель значительной части яиц. Также возрастает число различных аномалий развития и уродств. Интересно, что яйца одной кладки одной и той же особи имеют, по-видимому, разную степень сопротивляемости по отношению к вредящим факторам. Одни яйца гибнут, другие дают двойни или уродства, а третьи развиваются нормально. Сущность этих различий еще недостаточно выяснена. Есть также видовые различия реакции на вредные факторы. Форель, например, дает заметно больший процент близнецов, чем фундулус. По-видимому, в яйцах форели легче возникает физиологическая изоляция отдельных участков под влиянием вредящих воздействий, чем в яйцах фундулуса, согласно представлению Стокарда. В зависимости от того, на какой стадии эмбриогенеза действовал вредящий фактор, происходит остановка развития тех органов, которые наиболее интенсивно развиваются на этой стадии, а в связи с этим — соответствующие дефекты развития этих органов (мозга, глаз, конечностей и т. д.). Вредящее, искажающее развитие влияние оказывает также на своего партнера более развитой компонент, автозит на паразита, который в свою очередь тоже не может быть безразличным для автозита.
Другая серия опытов с целью получения близнецов была предпринята на яйцах фундулуса, подвергавшихся минутному облучению ультрафиолетовыми лучами (Hinrichs a. Genther, 1931). Яйца облучались, начиная с 5 минут после оплодотворения. Наибольшее число близнецов получалось под влиянием облучения на 25—30-й минуте и достигало 8.9%. И в этих опытах образование близнецов сопровождалось множественной гибелью яиц и разнообразными уродствами. Облучение за первые полчаса после оплодотворения яиц приходилось на такой период их жизни, когда дробление еще не начиналось или только что начиналось и никакой видимой морфологической дифференцировки еще не было. Большинство близнецов возникало, несомненно,из одного бластодиска, разделившегося, как думают авторы, еще задолго до гаструляции. Образование двух бластодисков на одном яйце с достоверностью не установлено. Раннее облучение способствует раздвоению переднего конца эмбриона, более позднее — заднего, поскольку развитие начинается с головного конца. Интересны некоторые числовые отношения полученных близнецов при облучении не позже, чем через 20 минут после оплодотворения; например, из 106 близнецов было получено: 5 пар с нормальными головами, 21 пара с одной нормальной и одной ненормальной головой, 9 пар, состоящих из слабо дифференцированной массы, одна пара имела частично раздвоенное сердце и 70 пар имели признаки слияния по основной оси (удвоенные ряды сомитов[12] и т. п., — рис. 85). Большинство, таким образом, состояло из соединенных близнецов, получившихся, вероятно, путем слияния двух самостоятельно образовавшихся зачатков, расположенных сравнительно близко друг от друга на общем бластодиске. Гаструляция на противоположных сторонах диска, т. е. под углом 180°, встречается сравнительно редко (рис. 85, Л). Микроскопическое изучение близнецов фундулуса, возникших под влиянием облучения как свободных, так и соединенных, обнаружило зеркальное расположение органов (Hinrichs, 1938). Рисунки, к сожалению, в этой работе не даны, и описание очень кратко; процент s. i. не указан.
 Влияние облучения на столь ранней стадии развития яиц можно пытаться объяснить тем, что облучением нарушается основной физиологический градиент, а в связи с этим возникают новые, вторичные градиенты, которые и оказываются основой образования ОБ.
Существует еще один прием получения близнецов у рыб — это пересадка участка верхней губы бластопора, физиологически наиболее активного участка гаструлы, в какое-нибудь другое место бластодиска. Трансплантат индуцирует развитие второго зародыша, что зависит от ряда конкретных условий опыта (Luther, 1935; Oppenheimer, 1936, и др.). Этот метод был впервые применен Шпеманом на амфибиях, вскоре мы с ним и познакомимся подробнее.
Влияние облучения на столь ранней стадии развития яиц можно пытаться объяснить тем, что облучением нарушается основной физиологический градиент, а в связи с этим возникают новые, вторичные градиенты, которые и оказываются основой образования ОБ.
Существует еще один прием получения близнецов у рыб — это пересадка участка верхней губы бластопора, физиологически наиболее активного участка гаструлы, в какое-нибудь другое место бластодиска. Трансплантат индуцирует развитие второго зародыша, что зависит от ряда конкретных условий опыта (Luther, 1935; Oppenheimer, 1936, и др.). Этот метод был впервые применен Шпеманом на амфибиях, вскоре мы с ним и познакомимся подробнее.
Амфибии
В естественных условиях близнецы у амфибий образуются очень редко. У лягушек описаны двухголовые головастики — форма соединенных близнецов, и у амфибий встречающаяся чаще других. Интересно, что при всей редкости двойных уродств у лягушек в одной партии яиц было обнаружено 27 экземпляров двухголовых головастиков. Чем это вызвано, объяснить трудно (Schwind, 1942). В других случаях описаны только единичные двойные уродства у разных видов лягушек (Лебединский, 1923, и др.). В некоторых случаях близнецы у лягушек получаются из перезрелых яиц (Witsch, 1934). Также редки случаи «спонтанно» возникающих в природе соединенных близнецов у хвостатых амфибий — саламандр, амблистом, тритонов и других (Schwind, 1942, и др.). Экспериментально близнецы у амфибий получены многими исследователями и сравнительно хорошо изучены. Яйцо тритона и лягушки оказалось удобным для таких экспериментов. Оно, как известно, имеет относительно небольшой запас желтка, расположенного преимущественно у вегетативного полюса, в связи с чем у некоторых видов амфибий вегетативный полюс имеет беловатую окраску, тогда как анимальный — темную; это помогает различать полюса. В зависимости от указанного распределения желтка яйца большинства амфибии имеют полное, но неравномерное дробление — у вегетативного полюса получаются более крупные бластомеры, чем у анимального. Близнецы у амфибий были получены искусственно разными способами, например: брались оплодотворенные яйца лягушек, еще не начавшие дробление или только его начинающие, помещались между двумя стеклянными пластинками анимальным полюсом вверх, как обычно яйца лягушек держатся в воде, и затем вместе со стеклами переворачивались на 180°. Благодаря легкому сжатию между стеклами яйца не могли вернуться в нормальное положение и оставались обращенными вегетативным полюсом вверх. Более тяжелые, крупные зерна желтка вегетативного полюса постепенно перемещались вниз, что можно было заметить по изменению окраски полюсов. В связи с этим происходило коренное перераспределение вещества в яйце, задержка развития и его нарушение, сопровождавшееся различными аномалиями и образованием близнецов, — например возникала двойная гаструляция и другие разнообразные нарушения нормального развития (Schultze, 1894; Schleip, 1929, и др.). Часто возникали симметричные близнецы, как соединенные, так и свободные (рис. 86). Однако большинство погибало, не достигнув конца развития. Этим методом получены близнецы также у тритонов (Тонков, 1900, и др.). Более точный и совершенный метод был разработан Шпеманом и его школой (Spemann, 1901—1903, 1936 и др.). Это наиболее известный метод экспериментального получения близнецов у амфибий, краткое описание достигнутых им результатов можно найти даже в школьных учебниках. Задача заключалась в том, чтобы физически изолировать первые бластомеры яиц тритонов (школа Шпемана работала главным образом на них) по аналогии с тем, как изолировались бластомеры яиц морских ежей в опытах Дриша и других. Изоляция бластомеров яиц тритона удалась путем перетяжки их волосом, а также другими приемами. Разделяя яйцо тритона по первой борозде дробления, можно получить пару настоящих близнецов, каждый из которых развился из одного из первых двух бластомеров (рис. 87). Если намеренно оставить перемычку между бластомерами, то получаются соединенные в той или иной степени близнецы, например с передним раздвоением до середины тела (рис. 60). При этом правый компонент, как и у рыб, может оказаться с зеркальным расположением внутренних органов, с situs inversus.
Более точный и совершенный метод был разработан Шпеманом и его школой (Spemann, 1901—1903, 1936 и др.). Это наиболее известный метод экспериментального получения близнецов у амфибий, краткое описание достигнутых им результатов можно найти даже в школьных учебниках. Задача заключалась в том, чтобы физически изолировать первые бластомеры яиц тритонов (школа Шпемана работала главным образом на них) по аналогии с тем, как изолировались бластомеры яиц морских ежей в опытах Дриша и других. Изоляция бластомеров яиц тритона удалась путем перетяжки их волосом, а также другими приемами. Разделяя яйцо тритона по первой борозде дробления, можно получить пару настоящих близнецов, каждый из которых развился из одного из первых двух бластомеров (рис. 87). Если намеренно оставить перемычку между бластомерами, то получаются соединенные в той или иной степени близнецы, например с передним раздвоением до середины тела (рис. 60). При этом правый компонент, как и у рыб, может оказаться с зеркальным расположением внутренних органов, с situs inversus.
 Эти опыты можно различно видоизменять. Например, если перетяжкой вскоре после оплодотворения яйца отделить ту часть яйца, которая содержит ядро, от той, которая его не имеет, то первая часть будет дробиться, а вторая останется без дробления (рис. 88). Если затем в недробящуюся часть пропустить одно из ядер через перемычку из другой части, то эта недробившаяся часть, получив ядро, начнет тоже дробиться, но с соответствующим опозданием по сравнению с другой частью, в результате чего получатся близнецы с разной степенью зрелости.
Первая борозда дробления, по которой накладывается лигатура при получении ОБ, не всегда одинаково разделяет яйцо в смысле более тонкой структуры его вещества. Одна деталь ее обнаруживается дополнительно вскоре после оплодотворения яйца в виде появления в области экватора яйца сероватого цвета полоски, по форме напоминающей серп, почему она и называется «серым серпом». Он находится в той части яйца, которая впоследствии становится спиной зародыша. Первая борозда дробления может проходить различно в отношении серого серпа. Если она рассечет его пополам, и в этом случае произвести перетяжку, то получатся два равноценных близнеца. Если же первая борозда разделит яйцо так, что серый серп целиком попадет в один из бластомеров, то при перетяжке целый зародыш получится только из того бластомера, в котором имеется серый серп. Из другого же бластомера, лишенного серого серпа, получится шарообразный ком клеток, не способный к дальнейшей дифференцировке (рис. 89). Если благодаря ослаблению перетяжки между обоими бластомерами и их дальнейшими производными оставить перемычку, то ком клеток, получившийся из «брюшного» бластомера, окажется подобием опухоли на теле нормального эмбриона, паразитом на нем, как на автозите. Возможно, что в свете этих опытов можно будет понять возникновение некоторых опухолей (тератом) у высших животных и человека. Методика таких перетяжек была с успехом применена и на безхвостых амфибиях, у которых также можно получить различные формы близнецов (Шмидт, 1933, и др.).
Эти опыты можно различно видоизменять. Например, если перетяжкой вскоре после оплодотворения яйца отделить ту часть яйца, которая содержит ядро, от той, которая его не имеет, то первая часть будет дробиться, а вторая останется без дробления (рис. 88). Если затем в недробящуюся часть пропустить одно из ядер через перемычку из другой части, то эта недробившаяся часть, получив ядро, начнет тоже дробиться, но с соответствующим опозданием по сравнению с другой частью, в результате чего получатся близнецы с разной степенью зрелости.
Первая борозда дробления, по которой накладывается лигатура при получении ОБ, не всегда одинаково разделяет яйцо в смысле более тонкой структуры его вещества. Одна деталь ее обнаруживается дополнительно вскоре после оплодотворения яйца в виде появления в области экватора яйца сероватого цвета полоски, по форме напоминающей серп, почему она и называется «серым серпом». Он находится в той части яйца, которая впоследствии становится спиной зародыша. Первая борозда дробления может проходить различно в отношении серого серпа. Если она рассечет его пополам, и в этом случае произвести перетяжку, то получатся два равноценных близнеца. Если же первая борозда разделит яйцо так, что серый серп целиком попадет в один из бластомеров, то при перетяжке целый зародыш получится только из того бластомера, в котором имеется серый серп. Из другого же бластомера, лишенного серого серпа, получится шарообразный ком клеток, не способный к дальнейшей дифференцировке (рис. 89). Если благодаря ослаблению перетяжки между обоими бластомерами и их дальнейшими производными оставить перемычку, то ком клеток, получившийся из «брюшного» бластомера, окажется подобием опухоли на теле нормального эмбриона, паразитом на нем, как на автозите. Возможно, что в свете этих опытов можно будет понять возникновение некоторых опухолей (тератом) у высших животных и человека. Методика таких перетяжек была с успехом применена и на безхвостых амфибиях, у которых также можно получить различные формы близнецов (Шмидт, 1933, и др.).

 Шпеман и его школа разработали еще другой прием для изучения вопросов детерминации на ранних стадиях эмбрионов амфибий, который вместе с тем служит для получения однояйцевых близнецов. С помощью особой микропипетки пересаживается (трансплантируется) участок верхней губы бластопора, как известно, физиологически наиболее активной области гаструлы, на другое менее активное место зародыша, и там, под влиянием этого трансплантата («организатора» по Шпеману) развивается второй зародыш (рис. 90), обычно более слабый, оказывающийся своего рода паразитом на теле основного зародыша. Этот метод получения добавочных зародышей был с успехом применен и на других позвоночных: у рыб, как говорилось уже выше, и у птиц, о чем еще будет речь в соответственном месте.
Сложную механику развития соединенных близнецов в известной мере освещает особая серия опытов Шпемана (Speman, 1903) и его сотрудников (Wessel, 1926; Koether, 1927, и др.). Здесь берутся две половины гаструл разного цвета, именно спинные части с бластопором, и сращиваются. Если рост осевых органов направлен навстречу друг другу, то при встрече они мешают друг другу дальше расти вперед и, развиваясь, расходятся под прямым углом к осям туловища, образуя характерное крестовидное удвоение, duplicitas cruciata по Шпеману (рис. 91). Столкнувшиеся передние участки образуют голову и передний отдел туловища, причем голова имеет два «лица», смотрящие в противоположные стороны, как у Януса (рис. 92), и каждое из этих «лиц» получается путем слияния половины тканей одной первоначальной гаструлы и половины другой, что видно по разнице окраски сращенных полугаструл. Остальная часть туловища каждого из компонентов состоит из материала только одной из первоначальных полугаструл. Эти части тела так и остаются под прямым углом к «лицам» составной головы, иначе говоря, плоскость симметрии головы расположена под прямым углом к плоскости симметрии задних областей туловища (рис. 92). Этот интересный двойной урод, полученный экспериментально, аналогичен удивительным уродствам млекопитающих и человека,возникающим спонтанно (рис.47). Очевидно, что при естественном образовании такого урода надо предполагать возникновение двух пунктов гаструляции с ростом осевых органов навстречу друг другу, что в опыте с тритоном достигается искусственной ориентацией обеих полугаструл при их сращивании.
Шпеман и его школа разработали еще другой прием для изучения вопросов детерминации на ранних стадиях эмбрионов амфибий, который вместе с тем служит для получения однояйцевых близнецов. С помощью особой микропипетки пересаживается (трансплантируется) участок верхней губы бластопора, как известно, физиологически наиболее активной области гаструлы, на другое менее активное место зародыша, и там, под влиянием этого трансплантата («организатора» по Шпеману) развивается второй зародыш (рис. 90), обычно более слабый, оказывающийся своего рода паразитом на теле основного зародыша. Этот метод получения добавочных зародышей был с успехом применен и на других позвоночных: у рыб, как говорилось уже выше, и у птиц, о чем еще будет речь в соответственном месте.
Сложную механику развития соединенных близнецов в известной мере освещает особая серия опытов Шпемана (Speman, 1903) и его сотрудников (Wessel, 1926; Koether, 1927, и др.). Здесь берутся две половины гаструл разного цвета, именно спинные части с бластопором, и сращиваются. Если рост осевых органов направлен навстречу друг другу, то при встрече они мешают друг другу дальше расти вперед и, развиваясь, расходятся под прямым углом к осям туловища, образуя характерное крестовидное удвоение, duplicitas cruciata по Шпеману (рис. 91). Столкнувшиеся передние участки образуют голову и передний отдел туловища, причем голова имеет два «лица», смотрящие в противоположные стороны, как у Януса (рис. 92), и каждое из этих «лиц» получается путем слияния половины тканей одной первоначальной гаструлы и половины другой, что видно по разнице окраски сращенных полугаструл. Остальная часть туловища каждого из компонентов состоит из материала только одной из первоначальных полугаструл. Эти части тела так и остаются под прямым углом к «лицам» составной головы, иначе говоря, плоскость симметрии головы расположена под прямым углом к плоскости симметрии задних областей туловища (рис. 92). Этот интересный двойной урод, полученный экспериментально, аналогичен удивительным уродствам млекопитающих и человека,возникающим спонтанно (рис.47). Очевидно, что при естественном образовании такого урода надо предполагать возникновение двух пунктов гаструляции с ростом осевых органов навстречу друг другу, что в опыте с тритоном достигается искусственной ориентацией обеих полугаструл при их сращивании.
 Если такие же полугаструлы ориентировать так, что рост осевых органов пойдет навстречу друг другу не прямо, а под углом, то можно получить различные формы с общей головой и в той или иной мере общим передним участком туловища, но разными задними отделами туловища и разными хвостами, заднее раздвоение — duplicitas posterior. Очевидно, что, ориентируя основные оси так, чтобы рост шел не навстречу, а, наоборот, расходился под углом, можно получить переднее раздвоение — d. anterior. Возможно, что в ряде случаев обе эти формы раздвоения в природе так и возникают: получаются сначала две точки гаструляции, в которых осевые органы растут под углом друг к другу: в зависимости от угла и расстояния бластопоров это приводит к вторичному слиянию той или иной области тела близнецов (Koether, 1927).
Многие подробности этих интересных явлений искусственного получении свободных и соединенных близнецов у амфибий еще не исследованы. Мы этих вопросов здесь касаться не можем. Но, во всяком случае, эксперименты на тритонах значительно содействовали пониманию эмбриогенеза ОБ.
Если такие же полугаструлы ориентировать так, что рост осевых органов пойдет навстречу друг другу не прямо, а под углом, то можно получить различные формы с общей головой и в той или иной мере общим передним участком туловища, но разными задними отделами туловища и разными хвостами, заднее раздвоение — duplicitas posterior. Очевидно, что, ориентируя основные оси так, чтобы рост шел не навстречу, а, наоборот, расходился под углом, можно получить переднее раздвоение — d. anterior. Возможно, что в ряде случаев обе эти формы раздвоения в природе так и возникают: получаются сначала две точки гаструляции, в которых осевые органы растут под углом друг к другу: в зависимости от угла и расстояния бластопоров это приводит к вторичному слиянию той или иной области тела близнецов (Koether, 1927).
Многие подробности этих интересных явлений искусственного получении свободных и соединенных близнецов у амфибий еще не исследованы. Мы этих вопросов здесь касаться не можем. Но, во всяком случае, эксперименты на тритонах значительно содействовали пониманию эмбриогенеза ОБ.
Рептилии
В естественных условиях соединенные близнецы описаны у различных рептилий: змей, черепах и других (Caullery, 1945; Алекперов, 1954, и др.). Обнаружены даже трехголовые змеи (Gemmill, 1912). У черепах описаны различные формы соединенных близнецов (рис. 93), известные у других животных и человека; раздвоенные спереди (duplicitas anterior) в разных вариантах, соединенные брюхом типа торакопагов и другие (Hildebrand, 1938, и др.). Установлено, что у некоторых видов черепах, например морской цейлонской черепахи, откладывающей яйца на берегу, раздвоение зародыша происходит очень часто (Caullery, 1945). Нечто подобное найдено у пресноводных американских черепах, причем образование близнецов часто сочетается с различными недостатками развития (Newman, 1923). Возникновение двоен, как и прочих аномалий, вероятно, зависит от воздействий окружающей среды, прежде всего от суточных колебаний температуры, которым подвергаются плохо защищенные яйца. На яйцах некоторых рептилий сделаны интересные наблюдения, проливающие свет на время образования близнецов у них. Так, на яйцах одной змеи — ужа (Tropidonotus natrix) описан случай начального развития двоен на стадии бластодиска (Wetzel, 1900). Из яйцевода было извлечено яйцо, имеющее четыре центра дробления, расположенные на поверхности яйца нормального размера (рис. 94). Каждый из этих бластодисков был несколько меньше нормального; между собой они были почти равны и содержали каждый от 60—65 до 73 клеток. Причины, их вызвавшие, мало понятны. Возможно, что сначала было два бластодиска, в свою очередь снова раздвоившихся. Вероятно, что каждый из этих четырех центров развития превратился бы в самостоятельную гаструлу, т. е. возникла бы четверня в одном яйце.
У той же змеи среди нормальных яиц обнаружено еще два яйца с двойнями на стадии гаструлы, причем у одного из них гаструлы находились на разных ступенях развития.
Соединенные и свободные близнецы на ранних стадиях развития были описаны у некоторых форм ящериц (Kopsch, 1897; Тур, 1903б, и др.). По-видимому, и у рептилий близнецы закладываются на ранних стадиях эмбриогенеза — до гаструляции и во время нее.
На яйцах некоторых рептилий сделаны интересные наблюдения, проливающие свет на время образования близнецов у них. Так, на яйцах одной змеи — ужа (Tropidonotus natrix) описан случай начального развития двоен на стадии бластодиска (Wetzel, 1900). Из яйцевода было извлечено яйцо, имеющее четыре центра дробления, расположенные на поверхности яйца нормального размера (рис. 94). Каждый из этих бластодисков был несколько меньше нормального; между собой они были почти равны и содержали каждый от 60—65 до 73 клеток. Причины, их вызвавшие, мало понятны. Возможно, что сначала было два бластодиска, в свою очередь снова раздвоившихся. Вероятно, что каждый из этих четырех центров развития превратился бы в самостоятельную гаструлу, т. е. возникла бы четверня в одном яйце.
У той же змеи среди нормальных яиц обнаружено еще два яйца с двойнями на стадии гаструлы, причем у одного из них гаструлы находились на разных ступенях развития.
Соединенные и свободные близнецы на ранних стадиях развития были описаны у некоторых форм ящериц (Kopsch, 1897; Тур, 1903б, и др.). По-видимому, и у рептилий близнецы закладываются на ранних стадиях эмбриогенеза — до гаструляции и во время нее.
Птицы
Вылупившиеся из яйца близнецы, как свободные, так и соединенные, у птиц почти неизвестны. Как редкий случай надо рассматривать появление из одного яйца двух нормальных цыплят — свободных близнецов. Они имели меньший, чем нормальный вес (11 и 16 г), оба женского пола, похожей окраски, с одним общим желточным мешком (рис. 95; Nalbandov, 1942). Такие близнецы, по-видимому, являются однояйцевыми в точном смысле слова, в отличие от двуяйцевых, хотя и вылупившихся из одной скорлупы, в которой находилось два «желтка», т. е. два яйца. Не менее редки случаи двойных уродств, как например цыпленок с двумя парами ног (рис. 96), т. е. duplicitas posterior (Кузнецов, 1951), или двухголовый жаворонок, у которого раздвоение имелось впереди от нижнего шейного позвонка (Caullery, 1945), наподобие двуглавого орла на гербе Византии и императорской России. Любопытен среди таких двойных уродств случай соединенных гомологичными местами черепа уток (рис. 97), которых приводит Бэр (1845а), аналогизируя их с изучавшимся им случаем карниопагов-младенцев (см. главу шестую).

 В зародышевом состоянии различного рода близнецы у птиц встречаются чаще, но они обычно гибнут, не достигая конца эмбрионального развития. Со времени Вольфа и Бэра такие близнецы в насиженном курином яйце описывались многократно (Dareste, 1891; Митрофанов, 1895; Kaestner, 1898— 1907; Tannreuther, 1919; Newman, 1923—1940; Politzer, 1955, и др.).
Обращаясь к эмбриогенезу близнецов у птиц, надо помнить, что у них яйцо имеет большой запас желтка, и потому дробление у птиц неполное, дискоидальное, сходное в этом отношении с яйцами костистых рыб и рептилий.
Близнецы у птиц могут происходить по крайней мере тремя способами: первый — образование двух и более бластодерм в одном яйце, второй — двойная гаструляция, третий — продольное раздвоение первоначально одного эмбриона, происходящее в период гаструляции. Кроме того, при втором способе образования близнецов легко может произойти сближение и слияние, в той или иной мере значительное, между обоими близнецами — возникновение двойных уродств.
Существование нескольких пунктов развития, распад бластодермы на 2—3 обособленных участка, явление, по-видимому, очень редкое, мало изученное. Сюда относятся, очевидно, препараты, описанные Дарестом, Кестнером и другими авторами (рис. 98). Зародыши смежных бластодерм одного яйца могут находиться иногда на разных стадиях развития. Оси зародышей могут быть направлены в разные стороны, как бы совершенно независимо друг от друга. Это первый способ образования ОБ. Второй трудно резко отграничить от него. Обособившийся участок бластодермы в дальнейшем переходит к самостоятельной гаструляции. Но возможно, что в единой бластодерме только на стадии гаструляции возникают два очага развития, т. е. гаструляция начинается в двух пунктах, притом иногда не одновременно. Это можнопредположить для тех случаев, когда зародыши находятся в явно разной степени зрелости (рис. 99). В ряде случаев зародыши в период гаструляции располагаются симметрично и растут прямо навстречу друг другу или под углом (рис. 100, 101). Это иногда приводит их к столкновению головными концами, деформации и вторичному соединению соприкасающимися частями, т. е. к образованию уродов с задним раздвоением (duplicitas posterior). Встречный рост эмбрионов не всегда кончается уродством. Это возможно даже в случае тройни, где все три эмбриона расположены с известной правильностью, наподобие трех радиусов из общего центра, к которому они обращены головой (рис. 102).
В зародышевом состоянии различного рода близнецы у птиц встречаются чаще, но они обычно гибнут, не достигая конца эмбрионального развития. Со времени Вольфа и Бэра такие близнецы в насиженном курином яйце описывались многократно (Dareste, 1891; Митрофанов, 1895; Kaestner, 1898— 1907; Tannreuther, 1919; Newman, 1923—1940; Politzer, 1955, и др.).
Обращаясь к эмбриогенезу близнецов у птиц, надо помнить, что у них яйцо имеет большой запас желтка, и потому дробление у птиц неполное, дискоидальное, сходное в этом отношении с яйцами костистых рыб и рептилий.
Близнецы у птиц могут происходить по крайней мере тремя способами: первый — образование двух и более бластодерм в одном яйце, второй — двойная гаструляция, третий — продольное раздвоение первоначально одного эмбриона, происходящее в период гаструляции. Кроме того, при втором способе образования близнецов легко может произойти сближение и слияние, в той или иной мере значительное, между обоими близнецами — возникновение двойных уродств.
Существование нескольких пунктов развития, распад бластодермы на 2—3 обособленных участка, явление, по-видимому, очень редкое, мало изученное. Сюда относятся, очевидно, препараты, описанные Дарестом, Кестнером и другими авторами (рис. 98). Зародыши смежных бластодерм одного яйца могут находиться иногда на разных стадиях развития. Оси зародышей могут быть направлены в разные стороны, как бы совершенно независимо друг от друга. Это первый способ образования ОБ. Второй трудно резко отграничить от него. Обособившийся участок бластодермы в дальнейшем переходит к самостоятельной гаструляции. Но возможно, что в единой бластодерме только на стадии гаструляции возникают два очага развития, т. е. гаструляция начинается в двух пунктах, притом иногда не одновременно. Это можнопредположить для тех случаев, когда зародыши находятся в явно разной степени зрелости (рис. 99). В ряде случаев зародыши в период гаструляции располагаются симметрично и растут прямо навстречу друг другу или под углом (рис. 100, 101). Это иногда приводит их к столкновению головными концами, деформации и вторичному соединению соприкасающимися частями, т. е. к образованию уродов с задним раздвоением (duplicitas posterior). Встречный рост эмбрионов не всегда кончается уродством. Это возможно даже в случае тройни, где все три эмбриона расположены с известной правильностью, наподобие трех радиусов из общего центра, к которому они обращены головой (рис. 102).


 Иногда трудно окончательно решить, как возникло заднее раздвоение. Например, рассмотрим случай, изображенный на рис.103. Таннрейтер (Tannreuther, 1919), описавший его, считает, что этот урод произошел путем слияния двух независимо друг от друга возникших первичных полосок, т. е. по второму из вышеназванных типов образования близнецов. Ньюмен (Newman, 1923, 1940а) не согласен С таким объяснением. Ссылаясь на полное отсутствие каких-либо следов слияния на общем переднем конце тела урода и полной симметрии правой и левой сторон этого участка, Ньюмен считает, что этот случай надо понимать, как результат продольного расщепления нижнего конца одного эмбриона; такой способ образования близнецов является, по мнению Ньюмена, самым распространенным. В доказательство существования этого способа он приводит другие случаи, например изображенный на рис.104, где два близнеца только в конце тела имеют небольшую перемычку и производят впечатление, будто бы фиксация препарата застигла их в конце процесса раздвоения, начавшегося с переднего конца. Другой пример Ньюмена (а их несколько) — это близнецы с параллельными осями (рис. 105) и т. д. Уоддингтон (Waddington, 1941) предполагает, что только спереди раздвоенные уроды (d. anterior; рис. 104) произошли путем расщепления потока тканей в процессе гаструляции, тогда как случаи заднего раздвоения, подобные изображенному на рис. 103, являются результатом слияния, вопреки мнению Ньюмена.
Иногда трудно окончательно решить, как возникло заднее раздвоение. Например, рассмотрим случай, изображенный на рис.103. Таннрейтер (Tannreuther, 1919), описавший его, считает, что этот урод произошел путем слияния двух независимо друг от друга возникших первичных полосок, т. е. по второму из вышеназванных типов образования близнецов. Ньюмен (Newman, 1923, 1940а) не согласен С таким объяснением. Ссылаясь на полное отсутствие каких-либо следов слияния на общем переднем конце тела урода и полной симметрии правой и левой сторон этого участка, Ньюмен считает, что этот случай надо понимать, как результат продольного расщепления нижнего конца одного эмбриона; такой способ образования близнецов является, по мнению Ньюмена, самым распространенным. В доказательство существования этого способа он приводит другие случаи, например изображенный на рис.104, где два близнеца только в конце тела имеют небольшую перемычку и производят впечатление, будто бы фиксация препарата застигла их в конце процесса раздвоения, начавшегося с переднего конца. Другой пример Ньюмена (а их несколько) — это близнецы с параллельными осями (рис. 105) и т. д. Уоддингтон (Waddington, 1941) предполагает, что только спереди раздвоенные уроды (d. anterior; рис. 104) произошли путем расщепления потока тканей в процессе гаструляции, тогда как случаи заднего раздвоения, подобные изображенному на рис. 103, являются результатом слияния, вопреки мнению Ньюмена.
 Изучая процесс гаструляции в курином яйце с помощью стереокинематографии Грэпер (Graper, 1931) наблюдал своеобразное движение тканей во время инвагинации, т. е. перемещения тканей в бластопор при образовании внутреннего слоя зародыша — энтодермы. Схематизируя возможные комбинации тканевых потоков при двойной гаструляции, в зависимость от которой Грэпер ставит возникновение близнецов, он старался «единообразно» объяснить образование различных типов соединенных близнецов (см. систему Швальбе, стр. 73). Схема Грэпера в наше время, однако, утратила свою универсальность и может годиться только для объяснения некоторых случаев (ср. Politzer, 1955).
Чем вызвано образование близнецов у птиц? Еще Дарест (Dareste, 1891), а позже Стокард (Stockard, 1921) и Ньюмен (Newman, 1923) высказали предположение, что близнецы возникают в тех яйцах, которые оказываются снесенными до окончания «критического» периода, когда еще возможно образование близнецов. Предполагается, что снесенное яйцо подвергается резкому охлаждению, которое вызывает задержку развития на некоторое время — это и способствует возникновению двух пунктов развития в силу их физиологической изоляции. Это то же самое объяснение, с которым мы уже встречались, касаясь образования близнецов у морских звезд, рыб, лягушек и т. д. Если допустить, что охлаждение застает яйцо на сравнительно поздней стадии развития — гаструляции, то вероятнее всего ожидать, что один, уже образовавшийся эмбрион подвергается расщеплению, т. е. близнецы чаще всего будут возникать по третьему способу, что, по мнению Ньюмена, и имеет место, притом с раздвоением заднего конца чаще, чем переднего, так как задний конец формируется позже переднего. В связи с этой гипотезой были поставлены опыты охлаждения яиц голубей, извлеченных досрочно из тела птицы (Riddle, 1923). Эти опыты не подтвердили гипотезы. Другая попытка проверки ее заключалась в охлаждении яиц до окончания гаструляции путем охлаждения самой птицы (курицы) в период начального развития в ней оплодотворенных яиц, до их кладки (Sturkie, 1946). На 73 обследованных так охлажденных яйцах было обнаружено 6 двоен, т. е. 8.2%, тогда как в норме двоен бывает около 0.111% (Byerly a. Olsen, 1934), т. e. увеличение близнецов на 8.09%, цифра статистически достоверная. Все экспериментальные эмбрионы отставали в развитии, из них 32.9% имели аномалии развития, не считая раздвоения. Близнецы были разных типов, все соединенные; малое число двоен не дает возможности сколько-нибудь основательно судить о численном отношении типов двойных уродств. Однако этими данными нельзя считать гипотезу Стокарда—Ньюмена доказанной, нужны дальнейшие опыты для ее проверки.
Близнецов у птиц вызывали и другими экспериментальными методами. Пользуясь методом Шпемана, трансплантировали кусочки различных тканей, могущих индуцировать образование в бластодерме добавочного зародыша — искусственного близнеца (Morita, 1936; Waddington, 1941). Эти явления аналогичны описанным у амфибий.
Путем механической изоляции — поперечным разрезом скальпелем — можно вызвать образование близнецов у утиного эмбриона (Lutz, 1953). Ориентация головы переднего партнера зависит от стадии гаструляции. Если разрез пройдет через участок, куда уже проникла образующаяся энтодерма (вспомним, что этот процесс идет от заднего конца будущего зародыша к переднему), то голова переднего близнеца будет ориентирована нормально, как и голова заднего, т. е. в одном направлении. Однако голова переднего эмбриона окажется ориентированной назад, навстречу голове заднего, если разрез провести на той стадии, когда энтодерма еще не образовалась в переднем участке. Это Лутц объясняет тем, что в этом последнем случае образование энтодермы потечет от переднего края бороздки, т. е. в обратном направлении, и потому голова переднего близнеца образуется у линии разреза, иначе говоря, будет обращена к голове заднего близнеца. Таким образом можно произвольно изменять относительное направление осей обоих близнецов.
Близнецов у птиц можно получить еще и другим приемом. На стадии ранней гаструляции, пока первичная полоска составляет одну четверть светлого поля бластодиска (area pellucida), наносится повреждение с помощью ультрафиолетового укола (по методу Чахотина) или электротока. В результате может возникнуть раздвоение процесса гаструляции и образование двоен, а изредка — троен. После того как первичная полоска достигла половины своей длины, близнецы этим методом уже не получаются (Twisselmann, 1939). Различные варианты получаемых близнецов зависели от конкретных условий опытов.
Таким образом, и стадия эмбриогенеза — не позже гаструляции, и приемы, вызывающие экспериментальное образование ОБ у птиц, в общем, аналогичны таковым у амфибий. Это говорит о сходстве по существу тех процессов, которые лежат в основе полиэмбрионии у этих животных.
Изучая процесс гаструляции в курином яйце с помощью стереокинематографии Грэпер (Graper, 1931) наблюдал своеобразное движение тканей во время инвагинации, т. е. перемещения тканей в бластопор при образовании внутреннего слоя зародыша — энтодермы. Схематизируя возможные комбинации тканевых потоков при двойной гаструляции, в зависимость от которой Грэпер ставит возникновение близнецов, он старался «единообразно» объяснить образование различных типов соединенных близнецов (см. систему Швальбе, стр. 73). Схема Грэпера в наше время, однако, утратила свою универсальность и может годиться только для объяснения некоторых случаев (ср. Politzer, 1955).
Чем вызвано образование близнецов у птиц? Еще Дарест (Dareste, 1891), а позже Стокард (Stockard, 1921) и Ньюмен (Newman, 1923) высказали предположение, что близнецы возникают в тех яйцах, которые оказываются снесенными до окончания «критического» периода, когда еще возможно образование близнецов. Предполагается, что снесенное яйцо подвергается резкому охлаждению, которое вызывает задержку развития на некоторое время — это и способствует возникновению двух пунктов развития в силу их физиологической изоляции. Это то же самое объяснение, с которым мы уже встречались, касаясь образования близнецов у морских звезд, рыб, лягушек и т. д. Если допустить, что охлаждение застает яйцо на сравнительно поздней стадии развития — гаструляции, то вероятнее всего ожидать, что один, уже образовавшийся эмбрион подвергается расщеплению, т. е. близнецы чаще всего будут возникать по третьему способу, что, по мнению Ньюмена, и имеет место, притом с раздвоением заднего конца чаще, чем переднего, так как задний конец формируется позже переднего. В связи с этой гипотезой были поставлены опыты охлаждения яиц голубей, извлеченных досрочно из тела птицы (Riddle, 1923). Эти опыты не подтвердили гипотезы. Другая попытка проверки ее заключалась в охлаждении яиц до окончания гаструляции путем охлаждения самой птицы (курицы) в период начального развития в ней оплодотворенных яиц, до их кладки (Sturkie, 1946). На 73 обследованных так охлажденных яйцах было обнаружено 6 двоен, т. е. 8.2%, тогда как в норме двоен бывает около 0.111% (Byerly a. Olsen, 1934), т. e. увеличение близнецов на 8.09%, цифра статистически достоверная. Все экспериментальные эмбрионы отставали в развитии, из них 32.9% имели аномалии развития, не считая раздвоения. Близнецы были разных типов, все соединенные; малое число двоен не дает возможности сколько-нибудь основательно судить о численном отношении типов двойных уродств. Однако этими данными нельзя считать гипотезу Стокарда—Ньюмена доказанной, нужны дальнейшие опыты для ее проверки.
Близнецов у птиц вызывали и другими экспериментальными методами. Пользуясь методом Шпемана, трансплантировали кусочки различных тканей, могущих индуцировать образование в бластодерме добавочного зародыша — искусственного близнеца (Morita, 1936; Waddington, 1941). Эти явления аналогичны описанным у амфибий.
Путем механической изоляции — поперечным разрезом скальпелем — можно вызвать образование близнецов у утиного эмбриона (Lutz, 1953). Ориентация головы переднего партнера зависит от стадии гаструляции. Если разрез пройдет через участок, куда уже проникла образующаяся энтодерма (вспомним, что этот процесс идет от заднего конца будущего зародыша к переднему), то голова переднего близнеца будет ориентирована нормально, как и голова заднего, т. е. в одном направлении. Однако голова переднего эмбриона окажется ориентированной назад, навстречу голове заднего, если разрез провести на той стадии, когда энтодерма еще не образовалась в переднем участке. Это Лутц объясняет тем, что в этом последнем случае образование энтодермы потечет от переднего края бороздки, т. е. в обратном направлении, и потому голова переднего близнеца образуется у линии разреза, иначе говоря, будет обращена к голове заднего близнеца. Таким образом можно произвольно изменять относительное направление осей обоих близнецов.
Близнецов у птиц можно получить еще и другим приемом. На стадии ранней гаструляции, пока первичная полоска составляет одну четверть светлого поля бластодиска (area pellucida), наносится повреждение с помощью ультрафиолетового укола (по методу Чахотина) или электротока. В результате может возникнуть раздвоение процесса гаструляции и образование двоен, а изредка — троен. После того как первичная полоска достигла половины своей длины, близнецы этим методом уже не получаются (Twisselmann, 1939). Различные варианты получаемых близнецов зависели от конкретных условий опытов.
Таким образом, и стадия эмбриогенеза — не позже гаструляции, и приемы, вызывающие экспериментальное образование ОБ у птиц, в общем, аналогичны таковым у амфибий. Это говорит о сходстве по существу тех процессов, которые лежат в основе полиэмбрионии у этих животных.
Млекопитающие
Среди этих животных есть два вида из отряда неполнозубых: броненосцы-армадилы (Dasypus novemcinctus и D. hybridus), которые замечательны тем, что закономерно размножаются путем образования нескольких зародышей из одного яйца (пол близнецов одного помета всегда одинаковый). Первый вид обычно имеет четырех однояйцевых близнецов, второй — разное число (до 12). Это явление полиэмбрионии у млекопитающих в известной мере аналогично вышеописанным случаям у кольчатых червей и паразитических насекомых. У млекопитающих это очень редкое явление. Другие виды броненосцев размножаются обычным путем, ОБ у них не описаны. У многих видов млекопитающих ОБ явление далеко не постоянное, наоборот, как мы увидим из дальнейшего, ОБ у остальных видов млекопитающих скорее исключение, чем правило. Оба вида броненосцев, имеющие ОБ, живут в Америке. Их удивительный способ размножения был давно известен. В науке он впервые описан в 1885 г. Иерингом, а подробно изучен только к начале XX в. Фернандец (Fernandez, 1909, 1913) изучил образование ОБ у южной формы (D. hybridus), а Ньюмен и Паттерсон (Newman а.Patterson, 1909; Patterson, 1913, 1927; Newman, 1923) — у техасского броненосца (D. novemcinctus). Этот последний был изучен подробнее, и потому мы его и рассмотрим в качестве примера. В пользу однояйцевости его близнецов прежде всего говорит тот факт, что возникновение их сопряжено с образованием в яичнике только одного желтого тела (Newman, 1923), что обычно связано с выходом одного яйца из фолликула, который и превращается в желтое тело яичника. Развитие яйца прослежено от поздних стадий дробления до полного развития близнецов (Patterson, 1913). В период половой активности, в августе, из оплодотворенного яйца начинает развиваться зародыш. В яйцеводах самки в это время можно найти самые ранние стадии эмбриогенеза, до бластоцисты включительно, т. е. до стадии полой сферы, ограниченной слоем клеток (трофобластом) с утолщением в одном участке — зародышевым узелком, из которого возникает сам зародыш (рис. 106).
 Бластоцнста, попав в матку, не сразу внедряется в ее стенку, а несколько недель остается неимплантированной. Это «период покоя», когда развитие, по-видимому, почти полностью останавливается. Осенью, после прикрепления к стенке матки, развитие бластоцисты возобновляется, при этом часть стенки бластоцисты превращается в плаценту, а в зародышевом узелке дифференцируются экто- и энтодерма (рис. 107). В разросшейся эктодерме развивается полость — это амниотическая полость, над ней возникает другая полость — внеамниотическая, и под амниотической полостью — желточный мешок; верхнюю стенку его образует энтодерма. На дне амниотической полости эктодерма утолщается — из нее в дальнейшем образуется зародышевый диск; это тот участок, где возникает сам зародыш. Полость амниона сплющивается и образует два боковых выроста, которые в дальнейшем в свою очередь раздваиваются. Так получается центральный участок амниотической полости и четыре ветви ее (рис. 108). В каждой из них затем возникает по первичной полоске, т. е. закладывается ось зародыша. Все оси, как лучи, обращены к общему центру, головным концом к нему (рис. 107, 3). В дальнейшем все четыре зародыша вытягиваются в длину, головы их обращены в противоположную от плаценты сторону. Таким образом, появлению зародышей близнецов предшествует своеобразное «почкование» амниотической полости с повторным удвоением. Несколько иначе протекает процесс разветвления амниотического пузыря у южного вида броненосца, где выросты («почки») возникают как бы беспорядочно, в разной последовательности, и оказываются различного размера и неодинаково расположенными по отношению друг к другу (рис. 109).
Бластоцнста, попав в матку, не сразу внедряется в ее стенку, а несколько недель остается неимплантированной. Это «период покоя», когда развитие, по-видимому, почти полностью останавливается. Осенью, после прикрепления к стенке матки, развитие бластоцисты возобновляется, при этом часть стенки бластоцисты превращается в плаценту, а в зародышевом узелке дифференцируются экто- и энтодерма (рис. 107). В разросшейся эктодерме развивается полость — это амниотическая полость, над ней возникает другая полость — внеамниотическая, и под амниотической полостью — желточный мешок; верхнюю стенку его образует энтодерма. На дне амниотической полости эктодерма утолщается — из нее в дальнейшем образуется зародышевый диск; это тот участок, где возникает сам зародыш. Полость амниона сплющивается и образует два боковых выроста, которые в дальнейшем в свою очередь раздваиваются. Так получается центральный участок амниотической полости и четыре ветви ее (рис. 108). В каждой из них затем возникает по первичной полоске, т. е. закладывается ось зародыша. Все оси, как лучи, обращены к общему центру, головным концом к нему (рис. 107, 3). В дальнейшем все четыре зародыша вытягиваются в длину, головы их обращены в противоположную от плаценты сторону. Таким образом, появлению зародышей близнецов предшествует своеобразное «почкование» амниотической полости с повторным удвоением. Несколько иначе протекает процесс разветвления амниотического пузыря у южного вида броненосца, где выросты («почки») возникают как бы беспорядочно, в разной последовательности, и оказываются различного размера и неодинаково расположенными по отношению друг к другу (рис. 109).

 Ньюмен считал, что «период покоя» бластоцисты до имплантации, сопровождающийся остановкой или задержкой развития, и есть причина образования близнецов у броненосцев, по аналогии с опытами Стокарда на рыбах. Против этого объяснения говорят многочисленные наблюдения на других млекопитающих — барсуках (Fischer, 1931), медведях (Verschuer, 1932), соболе (Наумов и Лавров, 1941),кунице (Enders a. Pearson, 1943), у которых бластоцисты также имеют «период покоя», однако ОБ у этих животных неизвестны. По-видимому, «почкование» амниотического пузыря у данных двух видов броненосцев есть не случайное явление, а нормальное видовое наследственное свойство. В реализации его «период покоя» может играть лишь роль одного из нужных условий (Gaullery. 1945).
Интересно отметить, что у армадилов образование выростов амниотической полости и раздвоение зародыша происходит в период, предшествующий закладке осевых органов, т. е. до гаструляции.
Открытие и исследование процесса образования ОБ у армадилов сыграло большую роль для понимания этого процесса у млекопитающих вообще, почему мы на нем и остановились подробнее.
Существует еще один род сумчатых млекопитающих — опоссумы, американские сумчатые крысы (Didelphys), у одного вида которых (D. virginiana) в левой матке была найдена четверня ОБ на ранней стадии развития. Расположение зародышей напоминало картину, описанную у армадила. Вместе с тем шесть бластоцист с одиночными эмбрионами обнаружены в правой матке того же самого животного (Patterson a. Hartman, 1917). Аналогичные наблюдения сделаны на другом виде опоссумов (D. marsupialis) еще в 1913 г. Полиэмбриония у этих животных носит спорадический характер, встречаясь наравне с обычным развитием яиц. Возможно, что таким путем вырабатывается регулярная полиэмбриония. У опоссумов этот вопрос систематически не изучался. Вообще относительно ОБ у диких животных имеется еще очень мало сведении. Встречаются лишь одиночные сообщения о редких казусах, как например о поимке двухголового дельфина у Антильских островов (Caullery, 1945), или отдельные изыскания в целях проверки гипотезы Ньюмена на барсуках, медведях и других животных.
Домашние животные в этом отношении изучены лучше, так же как и разводимые человеком для научных целей мыши, крысы, морские свинки, обезьяны и другие. К ним мы теперь и обратимся.
Ньюмен считал, что «период покоя» бластоцисты до имплантации, сопровождающийся остановкой или задержкой развития, и есть причина образования близнецов у броненосцев, по аналогии с опытами Стокарда на рыбах. Против этого объяснения говорят многочисленные наблюдения на других млекопитающих — барсуках (Fischer, 1931), медведях (Verschuer, 1932), соболе (Наумов и Лавров, 1941),кунице (Enders a. Pearson, 1943), у которых бластоцисты также имеют «период покоя», однако ОБ у этих животных неизвестны. По-видимому, «почкование» амниотического пузыря у данных двух видов броненосцев есть не случайное явление, а нормальное видовое наследственное свойство. В реализации его «период покоя» может играть лишь роль одного из нужных условий (Gaullery. 1945).
Интересно отметить, что у армадилов образование выростов амниотической полости и раздвоение зародыша происходит в период, предшествующий закладке осевых органов, т. е. до гаструляции.
Открытие и исследование процесса образования ОБ у армадилов сыграло большую роль для понимания этого процесса у млекопитающих вообще, почему мы на нем и остановились подробнее.
Существует еще один род сумчатых млекопитающих — опоссумы, американские сумчатые крысы (Didelphys), у одного вида которых (D. virginiana) в левой матке была найдена четверня ОБ на ранней стадии развития. Расположение зародышей напоминало картину, описанную у армадила. Вместе с тем шесть бластоцист с одиночными эмбрионами обнаружены в правой матке того же самого животного (Patterson a. Hartman, 1917). Аналогичные наблюдения сделаны на другом виде опоссумов (D. marsupialis) еще в 1913 г. Полиэмбриония у этих животных носит спорадический характер, встречаясь наравне с обычным развитием яиц. Возможно, что таким путем вырабатывается регулярная полиэмбриония. У опоссумов этот вопрос систематически не изучался. Вообще относительно ОБ у диких животных имеется еще очень мало сведении. Встречаются лишь одиночные сообщения о редких казусах, как например о поимке двухголового дельфина у Антильских островов (Caullery, 1945), или отдельные изыскания в целях проверки гипотезы Ньюмена на барсуках, медведях и других животных.
Домашние животные в этом отношении изучены лучше, так же как и разводимые человеком для научных целей мыши, крысы, морские свинки, обезьяны и другие. К ним мы теперь и обратимся.
Копытные. Корова обычно приносит одного теленка. Близнецы у коровы сравнительно редки, а ОБ среди них составляют меньший процент, чем у человека. Редкость случаев ОБ долго вызывала сомнение в возможности их появления у коров вообще. Существование ОБ у коров было с достоверностью установлено лишь благодаря трудам Кронахера (Kronacher, 1932) и его сотрудников, разработавших диагностику ОБ у коров по аналогии с созданной незадолго до того методике для различения двух типов близнецов у человека (см. глава четарнадцатая). Об этой методике Кронахера подробнее будет сказано дальше. По данным, собранным за последние три десятка лет при изучении свыше двухсот тысяч отелов, процент рождения близнецов колеблется в зависимости от материала (породы и проч.) от 4.61 до 1.00 и даже меньше (Bonnier a. Hansson, 1948; Gedda, 1951; Hancock, 1954). Процент ОБ среди близнецов не велик, приблизительно от 3 до 15 и больше при расчете по методу Вейнберга (см. главу девятую). Вычисление осложняется большей смертностью зародышей-близнецов мужского пола, чем женского (Hancock, 1954). Относительное количество рождающихся близнецов у одной и той же породы колеблется из года в год, как например у циммервальдского скота за 1906—1942 гг. (рис. 110; Weber, 1945). Причины этих колебаний недостаточно выяснены.
 Заметное влияние на многоплодие оказывает возраст коровы. С увеличением его идет наростание близнецов до известного предела, когда достигается максимум случаев рождения близнецов, а затем происходит перелом и резкий спад кривой (рис. 111). Это установлено различными исследователями (Johansson, 1932; Weber, 1945, и др.). Речь идет здесь о РБ. Аналогичные данные имеются и для человека (см. главу третью).
Другим фактором,от которого зависит многоплодие, является, по-видимому, наследственное предрасположение к нему. Установлены очень заметные различия в смысле склонности к рождению близнецов между отдельными породами коров, например в 5 раз больше у одной породы, чем у другой (Lush, 1925). Аналогичные различия найдены между отдельными линиями или «семьями» путем изучения родословных (Johansson, 1932; Lowe, 1939; Weber, 1945, и др.). Эти данные и выводы из них также аналогичны полученным у человека. Выступают семьи и особи, явно в большей мере предрасположенные к многоплодию, чем средняя масса. Например, одна корова последовательно принесла: двойню, одинца, двойню, одинца, пятерню — аборт на восьмом месяце (Caullery, 1945). Такое наследственное предрасположение идет не только по материнской линии, но и по отцовской (Weber, 1945, и др.). Установлены случаи, когда определенные быки у разных коров вызывали рождение близнецов, например от одного быка было 9 пар близнецов (Lush, 1925); подобные данные приводят и другие авторы (Johansson, 1932; Siems, 1933, и др.). В тех случаях, когда от такого быка рождаются ОБ, можно предположить, что сперма его стимулирует раздвоение зиготы или зародыша. Вообще же физиологический механизм влияния отца на рождение близнецов не понят до сих пор.
На увеличение многоплодия будто бы влияет характер кормления за определенный срок до оплодотворения (Wallace, 1952).
Введение гормона гипофиза увеличивает число созревающих одновременно яиц и тем самым число РБ, что впервые показал Завадовский (1941, 1946а, 1946б, 1954) и подтвердили другие авторы (Hammond, 1949). О методе Завадовского, используемого главным образом на овцах, подробнее сказано выше (см. главу третью). Здесь надо лишь отметить, что Завадовскому с помощью его метода удалось получать у коров не только двойни, но даже тройни, четверни и пятерни. Рождаемость телят в общем повышается от 101 —102 % к числу отелившихся коров до 120 — 140% (Завадовский, 1946б).
Близнецы в период эмбрионального развития оказывают нередко очень существенное влияние друг на друга благодаря близости имплантации в матке разных бластоцист и образованию анастомозов сосудов между плацентами (Swelt et al., 1940; Петской, 1953, и др.). Впервые Лилли (Lillie, 1917) объяснил, почему при рождении двух телят разного пола в ряде случаев телка оказывается в той или иной мере маскулинизированной, у нее в половых органах появляются признаки самца, а признаки ее пола, женского, оказываются редуцированными. Таких телок, как известно, стали называть «фри-мартины». Изменения эти происходят под влиянием мужского гормона, попадающего через кровь из мужского зародыша в женский благодаря анастомозам сосудов плацент (рис. 112). Предполагается, что мужская гонада развивается раньше женской, потому получаются только маскулинизированные самки, а обратное явление — феминизация самцов — не наблюдается. Фри-мартинов легко спутать в смысле пола, и это затрудняет в ряде случаев учет количества самцов и самок среди близнецов, который необходимо делать при применении метода Вейнберга для расчета числа ОБ. Кроме того, анастомозы сосудов, даже при наличии пары близнецов одинакового пола, способствуют большему сходству близнецов, чем это вызывается их генотипами, что может вести к затруднениям при диагнозе из принадлежности к ОБ или РБ (Swelt et al., 1940).
Заметное влияние на многоплодие оказывает возраст коровы. С увеличением его идет наростание близнецов до известного предела, когда достигается максимум случаев рождения близнецов, а затем происходит перелом и резкий спад кривой (рис. 111). Это установлено различными исследователями (Johansson, 1932; Weber, 1945, и др.). Речь идет здесь о РБ. Аналогичные данные имеются и для человека (см. главу третью).
Другим фактором,от которого зависит многоплодие, является, по-видимому, наследственное предрасположение к нему. Установлены очень заметные различия в смысле склонности к рождению близнецов между отдельными породами коров, например в 5 раз больше у одной породы, чем у другой (Lush, 1925). Аналогичные различия найдены между отдельными линиями или «семьями» путем изучения родословных (Johansson, 1932; Lowe, 1939; Weber, 1945, и др.). Эти данные и выводы из них также аналогичны полученным у человека. Выступают семьи и особи, явно в большей мере предрасположенные к многоплодию, чем средняя масса. Например, одна корова последовательно принесла: двойню, одинца, двойню, одинца, пятерню — аборт на восьмом месяце (Caullery, 1945). Такое наследственное предрасположение идет не только по материнской линии, но и по отцовской (Weber, 1945, и др.). Установлены случаи, когда определенные быки у разных коров вызывали рождение близнецов, например от одного быка было 9 пар близнецов (Lush, 1925); подобные данные приводят и другие авторы (Johansson, 1932; Siems, 1933, и др.). В тех случаях, когда от такого быка рождаются ОБ, можно предположить, что сперма его стимулирует раздвоение зиготы или зародыша. Вообще же физиологический механизм влияния отца на рождение близнецов не понят до сих пор.
На увеличение многоплодия будто бы влияет характер кормления за определенный срок до оплодотворения (Wallace, 1952).
Введение гормона гипофиза увеличивает число созревающих одновременно яиц и тем самым число РБ, что впервые показал Завадовский (1941, 1946а, 1946б, 1954) и подтвердили другие авторы (Hammond, 1949). О методе Завадовского, используемого главным образом на овцах, подробнее сказано выше (см. главу третью). Здесь надо лишь отметить, что Завадовскому с помощью его метода удалось получать у коров не только двойни, но даже тройни, четверни и пятерни. Рождаемость телят в общем повышается от 101 —102 % к числу отелившихся коров до 120 — 140% (Завадовский, 1946б).
Близнецы в период эмбрионального развития оказывают нередко очень существенное влияние друг на друга благодаря близости имплантации в матке разных бластоцист и образованию анастомозов сосудов между плацентами (Swelt et al., 1940; Петской, 1953, и др.). Впервые Лилли (Lillie, 1917) объяснил, почему при рождении двух телят разного пола в ряде случаев телка оказывается в той или иной мере маскулинизированной, у нее в половых органах появляются признаки самца, а признаки ее пола, женского, оказываются редуцированными. Таких телок, как известно, стали называть «фри-мартины». Изменения эти происходят под влиянием мужского гормона, попадающего через кровь из мужского зародыша в женский благодаря анастомозам сосудов плацент (рис. 112). Предполагается, что мужская гонада развивается раньше женской, потому получаются только маскулинизированные самки, а обратное явление — феминизация самцов — не наблюдается. Фри-мартинов легко спутать в смысле пола, и это затрудняет в ряде случаев учет количества самцов и самок среди близнецов, который необходимо делать при применении метода Вейнберга для расчета числа ОБ. Кроме того, анастомозы сосудов, даже при наличии пары близнецов одинакового пола, способствуют большему сходству близнецов, чем это вызывается их генотипами, что может вести к затруднениям при диагнозе из принадлежности к ОБ или РБ (Swelt et al., 1940).
 Недавно установлено еще одно интересное явление, зависящее от сообщения кровеносных систем близнецов, — это сходство реакций крови, несмотря на явную разнояйцевость зародышей, например у пятерни, состоящей из РБ. Речь идет о наследственных антигенах эритроцитов. Такая одинаковость реакции объясняется тем, что благодаря общности кровообращения у близнецов смешались те клетки, из которых впоследствии развились кроветворные органы; это вызвало своеобразный «мозаицизм» их. Явление такого рода у телят РБ уже неоднократно установлено (Owen, 1945; Owen et al., 1946). Это обстоятельство также может осложнить диагноз типа близнецов.
Многоплодие более высокого порядка, чем двойни, очень редкое явление у коров. Однако все же установлены случаи рождения троен и т. д. до шестерни включительно, причем некоторые из них однояйцевые (Johansson, 1932, Donald et al., 1951; Hancock, 1954, и др.). Число таких случаев мало, так что трудно делать какие-либо обобщения относительно закономерностей их состава.
Что касается соединенных близнецов, то они являются исключительной редкостью. Считается, что один такой случай приходится на 100 000 отелов (Keller a. Niedoba, 1937). Встречаются те же основные типы «сиамских близнецов», как у людей и других животных (рис. 113, 114). Интересно отметить, что в некоторых случаях имеется зеркальность в окраске шерсти обоих компонентов, когда один компонент выглядит как отражение другого в зеркале (рис. 115). Но в ряде случаев между компонентами имеются заметные различия и не только в окраске и распределении пятен на шерсти, но так же в извилинах головного мозга, в форме головы и т. д. (рис. 116, Keller u. Niedoba, 1937).
Недавно установлено еще одно интересное явление, зависящее от сообщения кровеносных систем близнецов, — это сходство реакций крови, несмотря на явную разнояйцевость зародышей, например у пятерни, состоящей из РБ. Речь идет о наследственных антигенах эритроцитов. Такая одинаковость реакции объясняется тем, что благодаря общности кровообращения у близнецов смешались те клетки, из которых впоследствии развились кроветворные органы; это вызвало своеобразный «мозаицизм» их. Явление такого рода у телят РБ уже неоднократно установлено (Owen, 1945; Owen et al., 1946). Это обстоятельство также может осложнить диагноз типа близнецов.
Многоплодие более высокого порядка, чем двойни, очень редкое явление у коров. Однако все же установлены случаи рождения троен и т. д. до шестерни включительно, причем некоторые из них однояйцевые (Johansson, 1932, Donald et al., 1951; Hancock, 1954, и др.). Число таких случаев мало, так что трудно делать какие-либо обобщения относительно закономерностей их состава.
Что касается соединенных близнецов, то они являются исключительной редкостью. Считается, что один такой случай приходится на 100 000 отелов (Keller a. Niedoba, 1937). Встречаются те же основные типы «сиамских близнецов», как у людей и других животных (рис. 113, 114). Интересно отметить, что в некоторых случаях имеется зеркальность в окраске шерсти обоих компонентов, когда один компонент выглядит как отражение другого в зеркале (рис. 115). Но в ряде случаев между компонентами имеются заметные различия и не только в окраске и распределении пятен на шерсти, но так же в извилинах головного мозга, в форме головы и т. д. (рис. 116, Keller u. Niedoba, 1937).




 Для использования ОБ коров в генетических целях как материала для близнецового метода весьма важным является правильная диагностика близнецов, различение ОБ и РБ. Для этой цели, как уже упоминалось, существует методика внутрипарного сравнения близнецов по некоторым наследственным признакам, которые у пары ОБ должны быть в значительной мере похожи, более похожи, чем у пары РБ. К таким признакам относятся форма и размеры некоторых частей тела, отпечаток кончика морды по аналогии с отпечатками пальцев у человека (рис. 117), завитки шерсти и другие особенности ее, цвет шерсти и расположение пятен на ней (рис. 118), пигментация кожи, некоторые особенности крови. Ни один из этих признаков не является решающим при диагнозе, и все они годятся для него с известными ограничениями. Критический разбор современного состояния диагноза типа близнецов у коров недавно опубликовал один из ведущих специалистов по близнецам у этих животных Хенкок (Hancock, 1954).
Интересно отметить, что была сделана попытка использовать для диагноза типа близнецов также пересадку кусочков кожи. Такие пересадки, как известно, сделанные с одной особи на другую, как правило, не удаются — пересаженный участок кожи погибает как чужеродное вещество. Успешными оказываются только пересадки на одном и том же индивидууме. Установлено, что у молодых коров пересадка кожи на разных степенях родства не удается. Только трансплантация с одного однояйцевого близнеца на другого той же пары дает положительный результат. Однако и с РБ происходит почти то же самое. Следовательно, для различения этих двух типов близнецов пересадки кожи не годятся. Но эти опыты возбудили вопрос — почему же удается пересадка у РБ? Предположительно можно лишь сказать, что у РБ благодаря анастомозам сосудов плацент может происходить смешение крови, вследствие чего возникает своеобразный «мозаицизм» крови по Оуэну и, вероятно, какая-то общность биохимии тканей, которая позволяет упомянутые пересадки (Anderson el. al., 1951). Возможно, что это явление сродни описанному Геддой (Gedda, 1951) сходству РБ по обмену (см. главу десятую «Органы кровообращения и кровь»). Только дальнейшее изучение этого интересного явления позволит объяснить его.
Применение близнецового метода, несмотря на относительно малое число близнецов у крупного рогатого скота, дало за последние годы ценные данные для суждения о роли наследственности в таких хозяйственно-важных свойствах коровы, как например различные свойства молока (содержание жира, казеина и т. д.), разумеется, при строго определенных условиях питания и содержания животных (Bonnier a. Hansson, 1946; King, 1953; Hancock, 1954, и др.). Это имеет большое значение для генетико-селекционной работы.
Изучается влияние количества и состава корма на увеличение роста (высота в плечах) и веса (живого веса) телят (Bonnier a. Hansson, 1948). Для этого одного из телят данной пары ОБ ставят на повышенный корм, другого — на пониженный. Снижение нормы питания не ниже известного критического лимита сказывается на весе, но не на росте. На последнем в большей мере, чем на весе, сказывается наследственная разница испытуемых пар (рис. 119, 120). Однако снижение рациона ниже критического лимита отзывается уже не только на весе, но и на росте (рис. 120). При статистической обработке такого материала специальными приемами получаются ценные выводы для животноводческой практики. В подробности мы здесь входить не можем (см. Hancock, 1954).
Для использования ОБ коров в генетических целях как материала для близнецового метода весьма важным является правильная диагностика близнецов, различение ОБ и РБ. Для этой цели, как уже упоминалось, существует методика внутрипарного сравнения близнецов по некоторым наследственным признакам, которые у пары ОБ должны быть в значительной мере похожи, более похожи, чем у пары РБ. К таким признакам относятся форма и размеры некоторых частей тела, отпечаток кончика морды по аналогии с отпечатками пальцев у человека (рис. 117), завитки шерсти и другие особенности ее, цвет шерсти и расположение пятен на ней (рис. 118), пигментация кожи, некоторые особенности крови. Ни один из этих признаков не является решающим при диагнозе, и все они годятся для него с известными ограничениями. Критический разбор современного состояния диагноза типа близнецов у коров недавно опубликовал один из ведущих специалистов по близнецам у этих животных Хенкок (Hancock, 1954).
Интересно отметить, что была сделана попытка использовать для диагноза типа близнецов также пересадку кусочков кожи. Такие пересадки, как известно, сделанные с одной особи на другую, как правило, не удаются — пересаженный участок кожи погибает как чужеродное вещество. Успешными оказываются только пересадки на одном и том же индивидууме. Установлено, что у молодых коров пересадка кожи на разных степенях родства не удается. Только трансплантация с одного однояйцевого близнеца на другого той же пары дает положительный результат. Однако и с РБ происходит почти то же самое. Следовательно, для различения этих двух типов близнецов пересадки кожи не годятся. Но эти опыты возбудили вопрос — почему же удается пересадка у РБ? Предположительно можно лишь сказать, что у РБ благодаря анастомозам сосудов плацент может происходить смешение крови, вследствие чего возникает своеобразный «мозаицизм» крови по Оуэну и, вероятно, какая-то общность биохимии тканей, которая позволяет упомянутые пересадки (Anderson el. al., 1951). Возможно, что это явление сродни описанному Геддой (Gedda, 1951) сходству РБ по обмену (см. главу десятую «Органы кровообращения и кровь»). Только дальнейшее изучение этого интересного явления позволит объяснить его.
Применение близнецового метода, несмотря на относительно малое число близнецов у крупного рогатого скота, дало за последние годы ценные данные для суждения о роли наследственности в таких хозяйственно-важных свойствах коровы, как например различные свойства молока (содержание жира, казеина и т. д.), разумеется, при строго определенных условиях питания и содержания животных (Bonnier a. Hansson, 1946; King, 1953; Hancock, 1954, и др.). Это имеет большое значение для генетико-селекционной работы.
Изучается влияние количества и состава корма на увеличение роста (высота в плечах) и веса (живого веса) телят (Bonnier a. Hansson, 1948). Для этого одного из телят данной пары ОБ ставят на повышенный корм, другого — на пониженный. Снижение нормы питания не ниже известного критического лимита сказывается на весе, но не на росте. На последнем в большей мере, чем на весе, сказывается наследственная разница испытуемых пар (рис. 119, 120). Однако снижение рациона ниже критического лимита отзывается уже не только на весе, но и на росте (рис. 120). При статистической обработке такого материала специальными приемами получаются ценные выводы для животноводческой практики. В подробности мы здесь входить не можем (см. Hancock, 1954).

 Обратимся к другим копытным домашним животным.
Многоплодие у овец более частое явление, чем у коров. В среднем, например, на 100 овец цигейской и каракульской породы приходится 140—150 развитых яиц (Завадовский, 1941). Количество созревающих яиц можно искусственно увеличить почти вдвое путем инъекции гормона гипофиза по методу Завадовского. Этим путем, конечно, увеличивается относительное число РБ. В естественных условиях относительное число ОБ, которое неоднократно устанавливалось у овец, составляет чуть ли не 20% всех пар одного пола (Gedda, 1951). Что касается распределения близнецов по полу, то получены цифры, близкие к ожидаемым теорией, например: 87 ММ: 184 МЖ : 90 ЖЖ (Chapman a. Lush, 1932) и другие в этом роде. Описаны различные случаи рождения более двух ягнят в один помет — до шести (Wilson a. Gregory, 1931). Шестерня состояла из трех самцов и трех самок. При применении метода Завадовского тоже рождались и пятерни, и шестерни. Как и у коров, у отдельных особей, а также некоторых пород наблюдается склонность к многоплодию, которая считается различными авторами наследственной (Завадовский, 1941; Caullery, 1945, и др.). Вопрос этот еще недостаточно изучен. Наблюдалось также влияние возраста овцы на многоплодие. Соединенные близнецы также не раз описывались у овец (Gedda, 1951).
Козы имеют еще большую склонность к многоплодию, чем овцы. Так, есть авторы, которые считают, что 75% пометов у коз — двойни (Wehefritz, 1925), причем нередки тройни и другие многоплодные пометы. Описаны и соединенные близнецы, что говорит о существовании у них ОБ, но последние еще мало изучены.
Свинья отличается значительным многоплодием. У нее в один помет бывает 6—10 поросят и даже свыше 20 (Wehefritz, 1925). ОБ встречаются, по-видимому, редко. Даже среди всех зародышей они составляют около 0.057% (Cohrs, 1934). Тем большую редкость представляет ранняя стадия развития ОБ, о которой речь будет ниже. Описаны также двойные уродства у свиней (Gedda, 1951), что свидетельствует о существовании у них ОБ.
Лошадь обычно приносит по одному жеребенку. В зависимости от породы рождается от 0.3 до 3% двоен. Указывается и на рождение троен. ОБ у лошади мало изучены; встречаются соединенные близнецы. По-видимому, ОБ у лошади очень редки (Caullery, 1945). Описаны лишь отдельные случаи ОБ (Uppenborn, 1933; Schermer, 1936, и др.).
У домашних хищников — собак и кошек, — размножающихся почти всегда пометом в несколько детенышей, которые обычно являются РБ, ОБ являются большой редкостью. В литературе имеется только один случай описания ОБ эмбрионов у собаки (Duke, 1946) и единичные случаи двойных уродств (Mainland, 1928, и др.). У кошки также описано всего несколько случаев соединенных близнецов, в XX в. — только один (рис. 121) — котенок с раздвоенной мордочкой (Bissonnette, 1933).
Обратимся к другим копытным домашним животным.
Многоплодие у овец более частое явление, чем у коров. В среднем, например, на 100 овец цигейской и каракульской породы приходится 140—150 развитых яиц (Завадовский, 1941). Количество созревающих яиц можно искусственно увеличить почти вдвое путем инъекции гормона гипофиза по методу Завадовского. Этим путем, конечно, увеличивается относительное число РБ. В естественных условиях относительное число ОБ, которое неоднократно устанавливалось у овец, составляет чуть ли не 20% всех пар одного пола (Gedda, 1951). Что касается распределения близнецов по полу, то получены цифры, близкие к ожидаемым теорией, например: 87 ММ: 184 МЖ : 90 ЖЖ (Chapman a. Lush, 1932) и другие в этом роде. Описаны различные случаи рождения более двух ягнят в один помет — до шести (Wilson a. Gregory, 1931). Шестерня состояла из трех самцов и трех самок. При применении метода Завадовского тоже рождались и пятерни, и шестерни. Как и у коров, у отдельных особей, а также некоторых пород наблюдается склонность к многоплодию, которая считается различными авторами наследственной (Завадовский, 1941; Caullery, 1945, и др.). Вопрос этот еще недостаточно изучен. Наблюдалось также влияние возраста овцы на многоплодие. Соединенные близнецы также не раз описывались у овец (Gedda, 1951).
Козы имеют еще большую склонность к многоплодию, чем овцы. Так, есть авторы, которые считают, что 75% пометов у коз — двойни (Wehefritz, 1925), причем нередки тройни и другие многоплодные пометы. Описаны и соединенные близнецы, что говорит о существовании у них ОБ, но последние еще мало изучены.
Свинья отличается значительным многоплодием. У нее в один помет бывает 6—10 поросят и даже свыше 20 (Wehefritz, 1925). ОБ встречаются, по-видимому, редко. Даже среди всех зародышей они составляют около 0.057% (Cohrs, 1934). Тем большую редкость представляет ранняя стадия развития ОБ, о которой речь будет ниже. Описаны также двойные уродства у свиней (Gedda, 1951), что свидетельствует о существовании у них ОБ.
Лошадь обычно приносит по одному жеребенку. В зависимости от породы рождается от 0.3 до 3% двоен. Указывается и на рождение троен. ОБ у лошади мало изучены; встречаются соединенные близнецы. По-видимому, ОБ у лошади очень редки (Caullery, 1945). Описаны лишь отдельные случаи ОБ (Uppenborn, 1933; Schermer, 1936, и др.).
У домашних хищников — собак и кошек, — размножающихся почти всегда пометом в несколько детенышей, которые обычно являются РБ, ОБ являются большой редкостью. В литературе имеется только один случай описания ОБ эмбрионов у собаки (Duke, 1946) и единичные случаи двойных уродств (Mainland, 1928, и др.). У кошки также описано всего несколько случаев соединенных близнецов, в XX в. — только один (рис. 121) — котенок с раздвоенной мордочкой (Bissonnette, 1933).

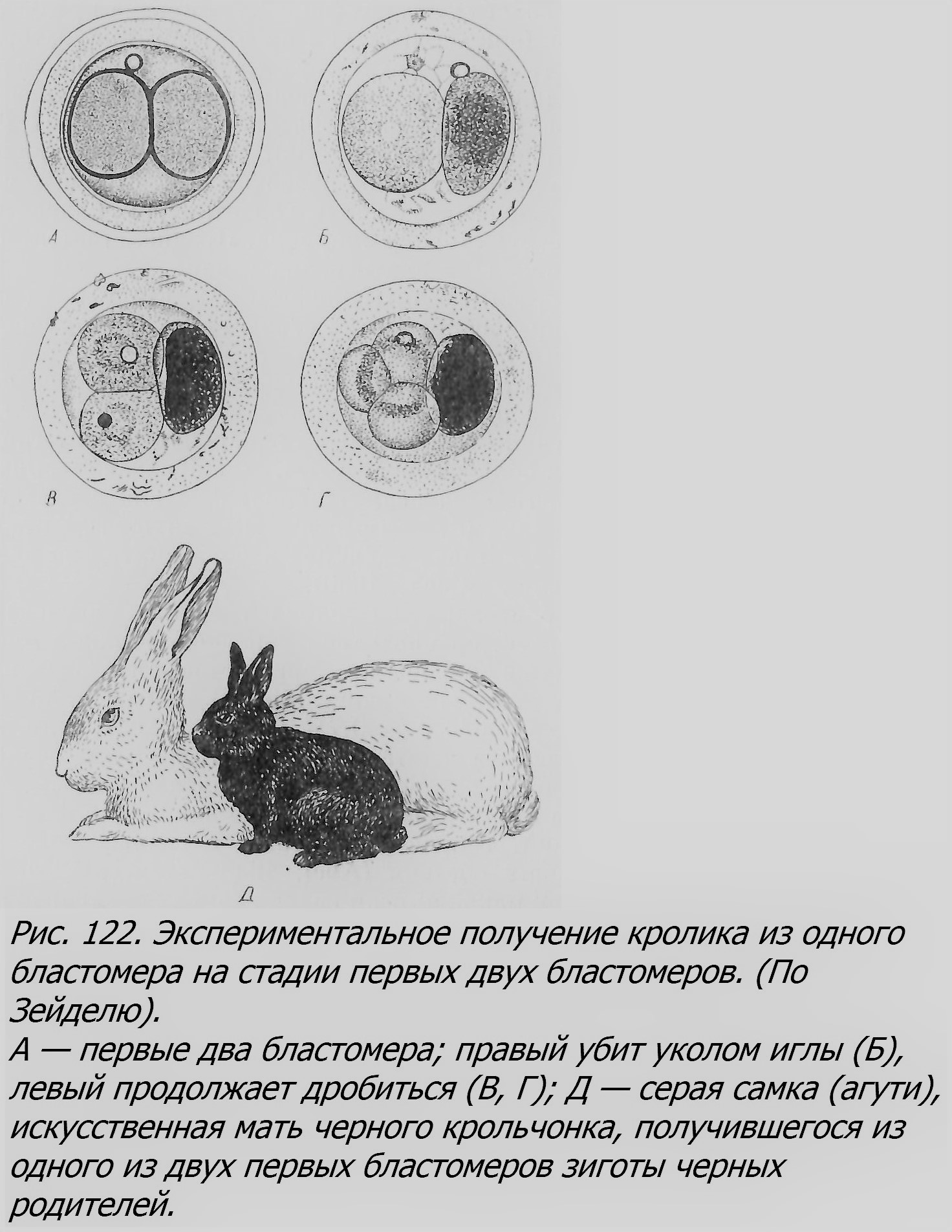 Грызуны, массами разводимые для экспериментальной работы — мыши, крысы, кролики, изучались и с точки зрения многоплодия. Все они имеют по нескольку детенышей в помете, обычно это РБ.
Кролики оказались удобным объектом для замечательного экспериментального показа возможности образования у них ОБ (Seidel, 1952). Извлеченные из яйцеводов зиготы кролика величиной 0.1 мм, на стадии двух первых бластомеров подвергались следующей! операции: уколом тонкой стеклянной иглы убивался один из двух бластомеров. Оставшийся бластомер мог продолжать жить и дробиться вне тела самки (рис. 122); убитый же бластомер отличался по цвету и малой прозрачности. Далее такие яйца, с продолжающим дробиться выжившим бластомером, переносились в соответственно подготовленную крольчиху. Из 27 таких «половинных» яйца два яйца закончили развитие, и у двух крольчих родилось по одному крольчонку, развившемуся из половины яйца. В пользу того, что эти крольчата именно такого происхождения, свидетельствует (кроме всех мер, которые гарантируют указанную беременность крольчих) еще тот факт, что окраска крольчат наследственно иная (они черные), чем их «матери»-инкубатора (агути). Интересно, что один из этих двух крольчат был вполне нормальный, тогда как другой имел некоторое искривление тела, напоминающее искривление тритонов, полученных Шпеманом путем перетяжки яйца (см. раздел «Амфибии»). Вероятно, искривление произошло в силу отставания развития той стороны зародыша, которая соприкасалась с убитым бластомером, повреждающе повлиявшим на эту сторону эмбриона. Предполагается, что крольчонок получился из правого бластомера.
Эти интересные факты показывают, что у млекопитающих возможно образование ОБ из первых двух бластомеров, так же как у других животных.
Сведений относительно ОБ у кроликов почти вовсе нет. Однако ОБ у них, по-видимому, все же возникают, так как описан случай соединенных близнецов (Fougeraux, 1786), являющийся большой редкостью. Лишь недавно описан новый случай заднего раздвоения, приблизительно с середины туловища, у кролика (d. posterior), с situs inversus у правого компонента и с некоторой разницей в форме пятна на раздвоенной части туловища (Cock, 1950).
Мыши, благодаря их плодовитости и наследственности окраски шерсти, сравнительно хорошо изученной генетически, могли бы служит удобным материалом для исследования вопросов близнечества. Но количество ОБ у мышей, по-видимому, очень мало — около 0.4—0.7% (Stewens, 1937, и др.). Описаны случаи двойных уродств. У крыс ОБ изучены очень мало.
В заключение остановимся на близнецах у обезьян, которых теперь тоже стали разводить в питомниках для опытных целей и наблюдений. Близнецы описаны у разных видов макак и других низших обезьян (Abel, 1933; Yerkes, 1934; Wislocki, 1939, и др.). У макак описан редкий случай двойного уродства — d. anterior. Животное имело две головы, двойной позвоночник, четыре руки, но один газ и одну пару нижних конечностей (Bolk, 1926). Другой случай меньшей степени переднего раздвоения описан тоже у макаки резуса (Hartman, 1943). Эти случаи свидетельствуют о существовании ОБ у обезьян, но они еще мало известны. У антропоидов (шимпанзе) описаны только РБ (Tomilin a. Yerkes, 1935, и др.). Всего до недавнего времени у обезьян установлено 24 случая близнецов. Они составляют приблизительно 1% всех родов, как у человека. Большинство близнецов — РБ (Schultz, 1956).
Грызуны, массами разводимые для экспериментальной работы — мыши, крысы, кролики, изучались и с точки зрения многоплодия. Все они имеют по нескольку детенышей в помете, обычно это РБ.
Кролики оказались удобным объектом для замечательного экспериментального показа возможности образования у них ОБ (Seidel, 1952). Извлеченные из яйцеводов зиготы кролика величиной 0.1 мм, на стадии двух первых бластомеров подвергались следующей! операции: уколом тонкой стеклянной иглы убивался один из двух бластомеров. Оставшийся бластомер мог продолжать жить и дробиться вне тела самки (рис. 122); убитый же бластомер отличался по цвету и малой прозрачности. Далее такие яйца, с продолжающим дробиться выжившим бластомером, переносились в соответственно подготовленную крольчиху. Из 27 таких «половинных» яйца два яйца закончили развитие, и у двух крольчих родилось по одному крольчонку, развившемуся из половины яйца. В пользу того, что эти крольчата именно такого происхождения, свидетельствует (кроме всех мер, которые гарантируют указанную беременность крольчих) еще тот факт, что окраска крольчат наследственно иная (они черные), чем их «матери»-инкубатора (агути). Интересно, что один из этих двух крольчат был вполне нормальный, тогда как другой имел некоторое искривление тела, напоминающее искривление тритонов, полученных Шпеманом путем перетяжки яйца (см. раздел «Амфибии»). Вероятно, искривление произошло в силу отставания развития той стороны зародыша, которая соприкасалась с убитым бластомером, повреждающе повлиявшим на эту сторону эмбриона. Предполагается, что крольчонок получился из правого бластомера.
Эти интересные факты показывают, что у млекопитающих возможно образование ОБ из первых двух бластомеров, так же как у других животных.
Сведений относительно ОБ у кроликов почти вовсе нет. Однако ОБ у них, по-видимому, все же возникают, так как описан случай соединенных близнецов (Fougeraux, 1786), являющийся большой редкостью. Лишь недавно описан новый случай заднего раздвоения, приблизительно с середины туловища, у кролика (d. posterior), с situs inversus у правого компонента и с некоторой разницей в форме пятна на раздвоенной части туловища (Cock, 1950).
Мыши, благодаря их плодовитости и наследственности окраски шерсти, сравнительно хорошо изученной генетически, могли бы служит удобным материалом для исследования вопросов близнечества. Но количество ОБ у мышей, по-видимому, очень мало — около 0.4—0.7% (Stewens, 1937, и др.). Описаны случаи двойных уродств. У крыс ОБ изучены очень мало.
В заключение остановимся на близнецах у обезьян, которых теперь тоже стали разводить в питомниках для опытных целей и наблюдений. Близнецы описаны у разных видов макак и других низших обезьян (Abel, 1933; Yerkes, 1934; Wislocki, 1939, и др.). У макак описан редкий случай двойного уродства — d. anterior. Животное имело две головы, двойной позвоночник, четыре руки, но один газ и одну пару нижних конечностей (Bolk, 1926). Другой случай меньшей степени переднего раздвоения описан тоже у макаки резуса (Hartman, 1943). Эти случаи свидетельствуют о существовании ОБ у обезьян, но они еще мало известны. У антропоидов (шимпанзе) описаны только РБ (Tomilin a. Yerkes, 1935, и др.). Всего до недавнего времени у обезьян установлено 24 случая близнецов. Они составляют приблизительно 1% всех родов, как у человека. Большинство близнецов — РБ (Schultz, 1956).
 Эмбриогенез близнецов, особенно ранний, у рассмотренных млекопитающих плохо изучен, так как материал очень редок. У овцы описана только одна бластоциста в возрасте около 7 дней с двумя зародышевыми узелками в период образования эндодермы (рис. 123), т. е. образования первичной полоски (Assheton, 1898, 1913). Другой случай ранней стадии развития ОБ в возрасте около 13 дней был описан у свиньи (Streeter, 1924). В матке одной свиньи среди нормальных яиц было обнаружено одно яйцо в стадии имплантации. Гистологическое изучение этого яйца показало, что оно уже прошло стадию бластоцисты и имело два расположенных друг против друга зародышевых диска с одним общим желточным мешком. Энтодерма была больше развита в области соприкосновения с каждым из зародышевых дисков (рис. 124). Один из зародышей (I) несколько отставал в развитии от другого (II): у последнего образование мезодермы было более выражено, чем у первого. Оба зародыша, по-видимому, находились на ранней стадии первичной полоски, т. е. в начале гаструляции. Стритер предполагает, что раздвоение яйца произошло во время образования бластоцисты: возникло два зародышевых узелка вместо одного. Чем это вызвано, остается неясным. У свиньи установлено вообще много аномалий раннего развития зиготы: около 10% яиц вообще не дробится, около 10% дегенерирует в стадии бластоцисты, от 5 до 10% развивается ненормально, и только около 70% превращается в жизнеспособных поросят. ОБ и в зародышевом состоянии у свиньи обнаруживаются редко. У других млекопитающих ранние стадии эмбриогенеза почти неизвестны. У облученных х-лучами мышей среди 1500 эмбрионов найден один случай ОБ, с общим желточным мешком, на стадии первичной полоски. Один из близнецов несколько более развит, чем другой. Оба были приблизительно на той же стадии развития, как и смежные одиночные эмбрионы. По-видимому, никакой задержки развития зиготы, из которой получились эти близнецы, не было; это противоречит гипотезе Ньюмена (Bodemann, 1935).
Мало материала имеется в распоряжении науки также относительно ранних стадий развития ОБ у человека. До недавнего времени был известен только один препарат на стадии первичной полоски (рис. 125), описанный Стритером (Streeter, 1919). Один из близнецов заметно опередил своего партнера, имеет амнион и желточный мешок. Длина этого зародыша 0.92 мм. Другой заметно меньше. Близнецы возникли, как предполагает Стритер, на стадии бластоцисты — образовалось два зародышевых узелка, подобно тому, как это показано на схеме Паттена (рис. 126).
Эмбриогенез близнецов, особенно ранний, у рассмотренных млекопитающих плохо изучен, так как материал очень редок. У овцы описана только одна бластоциста в возрасте около 7 дней с двумя зародышевыми узелками в период образования эндодермы (рис. 123), т. е. образования первичной полоски (Assheton, 1898, 1913). Другой случай ранней стадии развития ОБ в возрасте около 13 дней был описан у свиньи (Streeter, 1924). В матке одной свиньи среди нормальных яиц было обнаружено одно яйцо в стадии имплантации. Гистологическое изучение этого яйца показало, что оно уже прошло стадию бластоцисты и имело два расположенных друг против друга зародышевых диска с одним общим желточным мешком. Энтодерма была больше развита в области соприкосновения с каждым из зародышевых дисков (рис. 124). Один из зародышей (I) несколько отставал в развитии от другого (II): у последнего образование мезодермы было более выражено, чем у первого. Оба зародыша, по-видимому, находились на ранней стадии первичной полоски, т. е. в начале гаструляции. Стритер предполагает, что раздвоение яйца произошло во время образования бластоцисты: возникло два зародышевых узелка вместо одного. Чем это вызвано, остается неясным. У свиньи установлено вообще много аномалий раннего развития зиготы: около 10% яиц вообще не дробится, около 10% дегенерирует в стадии бластоцисты, от 5 до 10% развивается ненормально, и только около 70% превращается в жизнеспособных поросят. ОБ и в зародышевом состоянии у свиньи обнаруживаются редко. У других млекопитающих ранние стадии эмбриогенеза почти неизвестны. У облученных х-лучами мышей среди 1500 эмбрионов найден один случай ОБ, с общим желточным мешком, на стадии первичной полоски. Один из близнецов несколько более развит, чем другой. Оба были приблизительно на той же стадии развития, как и смежные одиночные эмбрионы. По-видимому, никакой задержки развития зиготы, из которой получились эти близнецы, не было; это противоречит гипотезе Ньюмена (Bodemann, 1935).
Мало материала имеется в распоряжении науки также относительно ранних стадий развития ОБ у человека. До недавнего времени был известен только один препарат на стадии первичной полоски (рис. 125), описанный Стритером (Streeter, 1919). Один из близнецов заметно опередил своего партнера, имеет амнион и желточный мешок. Длина этого зародыша 0.92 мм. Другой заметно меньше. Близнецы возникли, как предполагает Стритер, на стадии бластоцисты — образовалось два зародышевых узелка, подобно тому, как это показано на схеме Паттена (рис. 126).
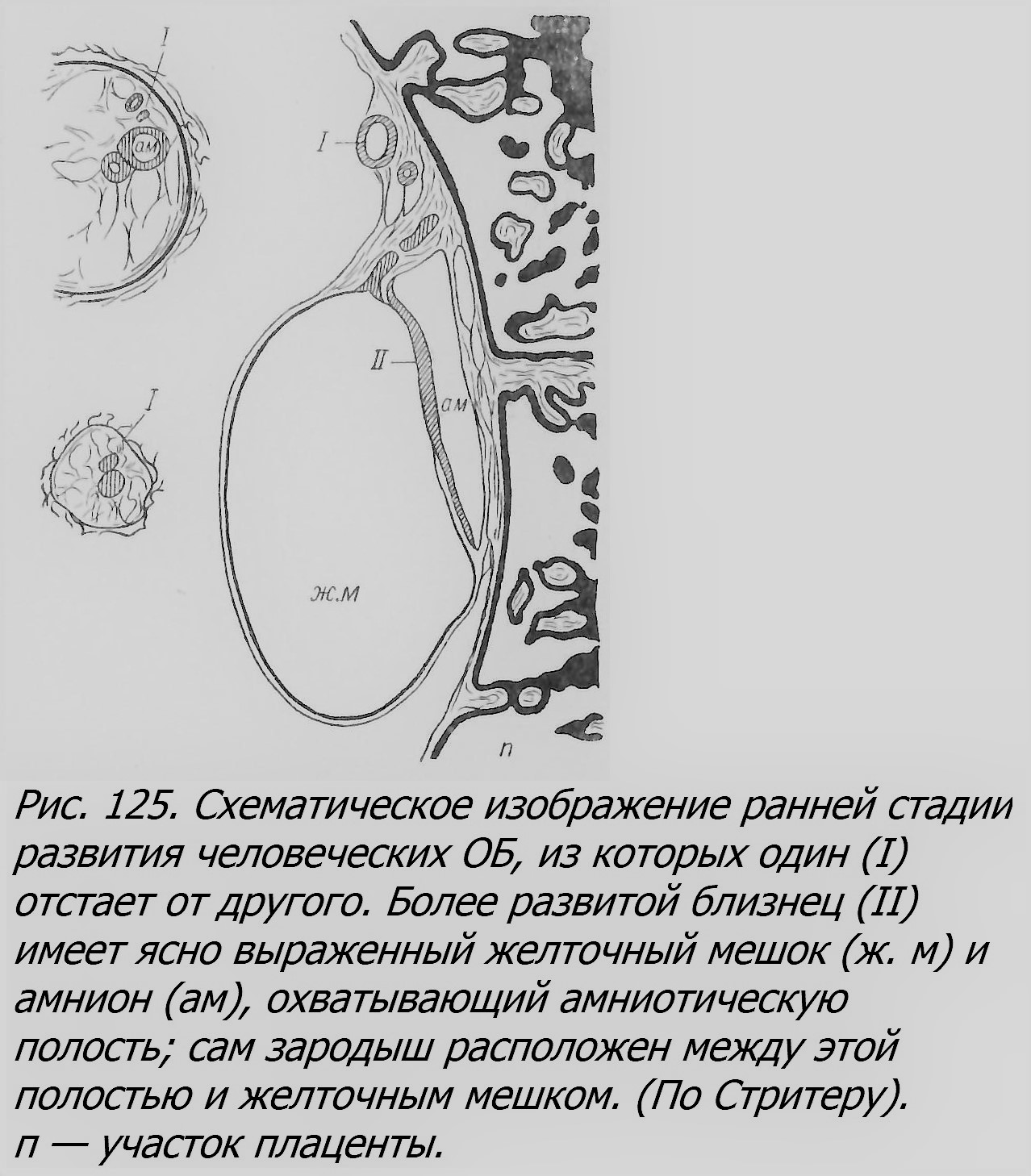
 Недавно описан еще один препарат человеческих ОБ на стадии первичной полоски, в возрасте около 17 дней (Corner et Baltimore, 1955). Эта пара считается самой юной из всех известных. Эти близнецы имеют общий хорион, но разные амнионы и желточные мешки. Другая пара, недавно описанная Мортоном (Morton, 1954), обнаруживает начальные стадии развития головы и конечностей. Этим зародышам близнецов около 32 дней. Они, как и предыдущая пара, имеют общий хорион и разные желточные мешки и амнионы (Corner et Baltimore, 1955). Из немногочисленных пар ОБ в возрасте 5—6 недель отметим еще пару монохориальных зародышей (рис. 126), одинаковой величины, длиной около 12 мм, в возрасте около 6 недель с общим желточным мешком (Агеу, 1922а). Возможно, что они возникли также на стадии бластоцисты, но можно предположить, что они произошли в результате двойной гаструляции (Newman, 1923).
Но вопросу об эмбриогенезе близнецов и, в частности, ОБ у человека на ранних стадиях развития эмбриона мы уже говорили в связи с вопросом о зародышевых оболочках и причинах возникновения близнецов (см. главу четвертую).
Весь рассмотренный небольшой материал по эмбриогенезу ОБ у млекопитающих, от броненосца до человека, показывает, что и у этого класса, как и у других животных, ОБ, очевидно, возникают тоже только на ранних стадиях развития: может быть, еще при первом дроблении зиготы, вероятно чаще всего до гаструляции, на стадии бластоцисты, и реже — на ранних этапах гаструляции.
Недавно описан еще один препарат человеческих ОБ на стадии первичной полоски, в возрасте около 17 дней (Corner et Baltimore, 1955). Эта пара считается самой юной из всех известных. Эти близнецы имеют общий хорион, но разные амнионы и желточные мешки. Другая пара, недавно описанная Мортоном (Morton, 1954), обнаруживает начальные стадии развития головы и конечностей. Этим зародышам близнецов около 32 дней. Они, как и предыдущая пара, имеют общий хорион и разные желточные мешки и амнионы (Corner et Baltimore, 1955). Из немногочисленных пар ОБ в возрасте 5—6 недель отметим еще пару монохориальных зародышей (рис. 126), одинаковой величины, длиной около 12 мм, в возрасте около 6 недель с общим желточным мешком (Агеу, 1922а). Возможно, что они возникли также на стадии бластоцисты, но можно предположить, что они произошли в результате двойной гаструляции (Newman, 1923).
Но вопросу об эмбриогенезе близнецов и, в частности, ОБ у человека на ранних стадиях развития эмбриона мы уже говорили в связи с вопросом о зародышевых оболочках и причинах возникновения близнецов (см. главу четвертую).
Весь рассмотренный небольшой материал по эмбриогенезу ОБ у млекопитающих, от броненосца до человека, показывает, что и у этого класса, как и у других животных, ОБ, очевидно, возникают тоже только на ранних стадиях развития: может быть, еще при первом дроблении зиготы, вероятно чаще всего до гаструляции, на стадии бластоцисты, и реже — на ранних этапах гаструляции.
Глава девятая КАК ЧАСТО РОЖДАЮТСЯ ЧЕЛОВЕЧЕСКИЕ БЛИЗНЕЦЫ?
Для многих видов животных многоплодие оказывается нормой, обычным явлением при размножении. Не пускаясь в рассмотрение всего многообразия этих феноменов, отметим лишь, что у различных видов млекопитающих самка, как правило, производит на свет несколько детенышей - обычно РБ, например у мышей, крыс, кроликов, кошек, собак, свиней и т. д. Рождение одного детеныша у таких животных исключение. Наоборот, для других видов это правило, а многоплодие исключение; сюда относятся корова, лошадь, слон и другие. До некоторой степени промежуточное положение в этом отношении занимают овцы, козы и другие животные. Для некоторых видов многоплодие, по-видимому, установившийся видовой признак, так же как для других таковым является одноплодие. И есть виды, у которых это свойство нечетко выражено, меняется в зависимости от породы, линии и т. д. О связи многоплодия с размерами тела вида или расы речь была выше (см. главу третью). Для человека нормой является беременность одним плодом, многоплодие представляет исключение. На основании большого материала различных стран можно считать, что приблизительно на 100 обычных родов приходятся одни роды близнецов, т. е. около 1%. Это, разумеется, цифра далеко не постоянная. У разных народов, в разные годы у одного и того же народа и т. д. процент рождения близнецов колеблется, то превышая 1.5, то падая ниже 0.5. Колебания эти пока еще недостаточно изучены, и причины их не вполне понятны. Частично такие колебания могут зависить и от техники статистического учета рождаемости близнецов, возникающих при этом неточностей: например, не всегда учитываются мертворожденные двойни, двойни с одним мертворожденным регистрируются как одиночные роды и т. д. Примером разницы в числе рождения близнецов в разных странах может служить таблица, составленная на основании изучения статистических данных относительно, примерно, 100 млн родов (рис. 127; Greulich, 1930). Однако порядок стран по относительному количеству близнецовых родов, по-видимому, не является постоянным. Так, по более поздним данным, Италия, например, стоит перед Германией и Болгарией, Венгрия стоит после США и т. д. (Gedda et al., 1955). Но и в разных районах одной страны частота близнецовых родов оказывается далеко не одинаковой. Так, например, в Италии в разных местностях частота разная: низшая 1.1 —1.2%, высшая 1.5—1.6% (рис. 128). Причины таких различий могут быть разнообразные: влияние широты и климата, расового состава, образа жизни (город или деревня) и экономики разных групп населения и т. д.
 Статистика рождаемости не отражает, как уже указывалось, реальной картины многоплодия у человека. Различные исследователи указывают на значительную смертность близнецов во время утробной жизни: часть их гибнет в виде спонтанных абортов, часть оказывается мертворожденными и т. д. Значительная часть гибнет вскоре после рождения, тем более что среди близнецов чаще встречаются недоноски, чем среди одиночек. Например, по американским данным за 1931—1936 гг., число мертворожденных и умерших вскоре после рождения близнецов более чем в 3 раза превышает смертность одиночек; среди близнецов одинакового пола смертность больше, чем среди близнецов разного пола, а среди ОБ вдвое больше, чем среди РБ. Около 30% родов близнецов преждевременные и т. д. (Yerushalmy a. Sheear, 1940b). Об этих вопросах уже говорилось выше.
До сих пор шла речь о частоте рождений близнецов вообще. Можно установить относительное количество рождений ОБ к рождению РБ. Это делается по методу Вейнберга (Weinberg, 1903,1928,1934). Поскольку РБ являются братьями и сестрами, рождающимися одновременно, надо допустить, что в смысле соотношения полов они, как и одиночки, должны быть приблизительно в обычном соотношении, т. е. поровну мужского и женского. Расчет (по теории вероятности) показывает, что пар одинакового пола, как того, так и другого, должно быть по 25%, а пар разного пола — 50%. Иначе говоря, в массе РБ половина пар должна быть неодинакового пола и равна числу пар РБ одинакового пола, т. е. всем РБ, где оба партнера или мужского, или женского пола. Это соотношение выражается так: 25% ММ+50% МЖ+25% ЖЖ, где М и Ж соответственно обозначают пол. Так как ОБ всегда составляют пары одинакового пола, то рассчитать их число можно очень просто: вычесть число пар разного пола, определить которые легко, из общего числа всех пар одинакового пола, как мужского, так и женского. Разность дает число пар ОБ. Но так как в действительности мальчиков рождается немного больше, чем девочек, то в расчет по методу Вейнберга вносится соответствующая поправка, которая мало меняет суть дела. Конкретным примером такого расчета могут служить цифры, полученные на основании статистических данных Германии (до второй мировой войны). На 85 родов приходилась одна пара близнецов вообще. Из них оказалось около 25% однояйцевых, что дает около 0.3% всех родов; иначе говоря, одна пара ОБ приходится приблизительно на 340 родов (Lotze, 1937).
Метод Вейнберга был проверен на населении Европы и США и признан различными исследователями (Dahlberg, 1926; Verschuer, 1932; Yerushalmy a. Sheear, 1940а; Newman, 1940с; Gedda, 1951; Allen, 1955b, и др.). Этим методом работают и в настоящее время. Оказалось, в разных странах число пар ОБ в общем составляет от 21 до 33.4% всех близнецов. В одной и той же стране, например в Италии, число ОБ в течение 5 лет (1931 —1935 гг.) колебалось от 28.5 до 30.2%, составляя в среднем 29.2%. А в Риме за 1938—1945 гг. среднее число пар ОБ составляло 32.93% с колебаниями в разные годы от 29.34 до 40,48% (Gedda, 1951). Мы видим, что отдельные пункты страны имеют и свою среднюю, и свои пределы колебаний в разные годы, отличные от общих цифр всей страны. Чем вызваны эти колебания, пока остается невыясненным.
Изучение близнецов в Японии обнаружило, что относительное число РБ здесь заметно меньше, чем в Европе, и составляет около 1/4—1/3 относительного числа РБ, встречающихся в населении европеоидов и негроидов; при этом относительное число японских ОБ в общем такое же, как и у этих рас. По-видимому, нечто подобное наблюдается и среди других народов монголоидной расы — в Корее, Индокитае и т. д. (Komai a. Fukuoka, 1936) и Южной Америке, там, где преобладает индейское население (Newman, 1940с), т. е. тоже монголоиды. Есть и другие указания на связь рождаемости близнецов с расой; например у северных народов Европы, имеющих светлые глаза, относительное число близнецов несколько больше, чем у южных (Dahlberg, 1926). Однако все эти сведения еще слишком недостаточны, чтобы их можно было обобщить и сделать какие-либо обоснованные выводы относительно зависимости многоплодия от расы у человека.
Надо еще отметить, что при учете относительного количества новорожденных РБ того и другого пола мальчиков оказывается меньше, чем при обычных родах. Относительное число мальчиков в общем убывает с увеличением числа одновременно рождающихся младенцев (Turpin et Schutzenberger, 1950, 1952а) (см. далее).
Теперь нам надо обратиться к вопросу об относительном числе более редких случаев многоплодия у людей: троен, чет-верен и т. д. Их частота определяется так называемым «законом Эллина» (Hellin, 1895). Согласно этому закону, многоплодные роды тем реже, чем больше число рождающихся одновременно младенцев, т. е. четверни родятся реже троен, и т. д. Это отношение имеет следующее численное выражение: если двойни рождаются в отношении 1 : 85, то тройни — в отношении 1 : 852, четверни — 1 : 853, и т. д. Иначе говоря, если число одновременно рождающихся будет «x», то шансы на такие роды будут 1 : 85(x-1). Закон Эллина неоднократно проверялся на больших цифрах статистического материала и в общем был подтвержден (Lotze, 1937; Strandskov, 1945; Gedda, 1951; Miettinen, 1954, и др.). Например, согласно немецкой статистике за 10 лет, двойни рождались приблизительно в количестве 1 : 85.6, тройни — 1 : 842, т. е. одна тройня приблизительно на 7000 родов, четверни — 1: 923, или одна четверня примерно на 780 000 родов. Это составляет около двух четверней в год на всю довоенную Германию. Соответственно пятерни и шестерни составляют еще большую редкость. Закон Эллина только приблизительно соответствует реальным фактам.Примером более детальным служит следующая таблица Гедды (Gedda, 1951), составленная на основании итальянской статистики (табл. 9).
Статистика рождаемости не отражает, как уже указывалось, реальной картины многоплодия у человека. Различные исследователи указывают на значительную смертность близнецов во время утробной жизни: часть их гибнет в виде спонтанных абортов, часть оказывается мертворожденными и т. д. Значительная часть гибнет вскоре после рождения, тем более что среди близнецов чаще встречаются недоноски, чем среди одиночек. Например, по американским данным за 1931—1936 гг., число мертворожденных и умерших вскоре после рождения близнецов более чем в 3 раза превышает смертность одиночек; среди близнецов одинакового пола смертность больше, чем среди близнецов разного пола, а среди ОБ вдвое больше, чем среди РБ. Около 30% родов близнецов преждевременные и т. д. (Yerushalmy a. Sheear, 1940b). Об этих вопросах уже говорилось выше.
До сих пор шла речь о частоте рождений близнецов вообще. Можно установить относительное количество рождений ОБ к рождению РБ. Это делается по методу Вейнберга (Weinberg, 1903,1928,1934). Поскольку РБ являются братьями и сестрами, рождающимися одновременно, надо допустить, что в смысле соотношения полов они, как и одиночки, должны быть приблизительно в обычном соотношении, т. е. поровну мужского и женского. Расчет (по теории вероятности) показывает, что пар одинакового пола, как того, так и другого, должно быть по 25%, а пар разного пола — 50%. Иначе говоря, в массе РБ половина пар должна быть неодинакового пола и равна числу пар РБ одинакового пола, т. е. всем РБ, где оба партнера или мужского, или женского пола. Это соотношение выражается так: 25% ММ+50% МЖ+25% ЖЖ, где М и Ж соответственно обозначают пол. Так как ОБ всегда составляют пары одинакового пола, то рассчитать их число можно очень просто: вычесть число пар разного пола, определить которые легко, из общего числа всех пар одинакового пола, как мужского, так и женского. Разность дает число пар ОБ. Но так как в действительности мальчиков рождается немного больше, чем девочек, то в расчет по методу Вейнберга вносится соответствующая поправка, которая мало меняет суть дела. Конкретным примером такого расчета могут служить цифры, полученные на основании статистических данных Германии (до второй мировой войны). На 85 родов приходилась одна пара близнецов вообще. Из них оказалось около 25% однояйцевых, что дает около 0.3% всех родов; иначе говоря, одна пара ОБ приходится приблизительно на 340 родов (Lotze, 1937).
Метод Вейнберга был проверен на населении Европы и США и признан различными исследователями (Dahlberg, 1926; Verschuer, 1932; Yerushalmy a. Sheear, 1940а; Newman, 1940с; Gedda, 1951; Allen, 1955b, и др.). Этим методом работают и в настоящее время. Оказалось, в разных странах число пар ОБ в общем составляет от 21 до 33.4% всех близнецов. В одной и той же стране, например в Италии, число ОБ в течение 5 лет (1931 —1935 гг.) колебалось от 28.5 до 30.2%, составляя в среднем 29.2%. А в Риме за 1938—1945 гг. среднее число пар ОБ составляло 32.93% с колебаниями в разные годы от 29.34 до 40,48% (Gedda, 1951). Мы видим, что отдельные пункты страны имеют и свою среднюю, и свои пределы колебаний в разные годы, отличные от общих цифр всей страны. Чем вызваны эти колебания, пока остается невыясненным.
Изучение близнецов в Японии обнаружило, что относительное число РБ здесь заметно меньше, чем в Европе, и составляет около 1/4—1/3 относительного числа РБ, встречающихся в населении европеоидов и негроидов; при этом относительное число японских ОБ в общем такое же, как и у этих рас. По-видимому, нечто подобное наблюдается и среди других народов монголоидной расы — в Корее, Индокитае и т. д. (Komai a. Fukuoka, 1936) и Южной Америке, там, где преобладает индейское население (Newman, 1940с), т. е. тоже монголоиды. Есть и другие указания на связь рождаемости близнецов с расой; например у северных народов Европы, имеющих светлые глаза, относительное число близнецов несколько больше, чем у южных (Dahlberg, 1926). Однако все эти сведения еще слишком недостаточны, чтобы их можно было обобщить и сделать какие-либо обоснованные выводы относительно зависимости многоплодия от расы у человека.
Надо еще отметить, что при учете относительного количества новорожденных РБ того и другого пола мальчиков оказывается меньше, чем при обычных родах. Относительное число мальчиков в общем убывает с увеличением числа одновременно рождающихся младенцев (Turpin et Schutzenberger, 1950, 1952а) (см. далее).
Теперь нам надо обратиться к вопросу об относительном числе более редких случаев многоплодия у людей: троен, чет-верен и т. д. Их частота определяется так называемым «законом Эллина» (Hellin, 1895). Согласно этому закону, многоплодные роды тем реже, чем больше число рождающихся одновременно младенцев, т. е. четверни родятся реже троен, и т. д. Это отношение имеет следующее численное выражение: если двойни рождаются в отношении 1 : 85, то тройни — в отношении 1 : 852, четверни — 1 : 853, и т. д. Иначе говоря, если число одновременно рождающихся будет «x», то шансы на такие роды будут 1 : 85(x-1). Закон Эллина неоднократно проверялся на больших цифрах статистического материала и в общем был подтвержден (Lotze, 1937; Strandskov, 1945; Gedda, 1951; Miettinen, 1954, и др.). Например, согласно немецкой статистике за 10 лет, двойни рождались приблизительно в количестве 1 : 85.6, тройни — 1 : 842, т. е. одна тройня приблизительно на 7000 родов, четверни — 1: 923, или одна четверня примерно на 780 000 родов. Это составляет около двух четверней в год на всю довоенную Германию. Соответственно пятерни и шестерни составляют еще большую редкость. Закон Эллина только приблизительно соответствует реальным фактам.Примером более детальным служит следующая таблица Гедды (Gedda, 1951), составленная на основании итальянской статистики (табл. 9).
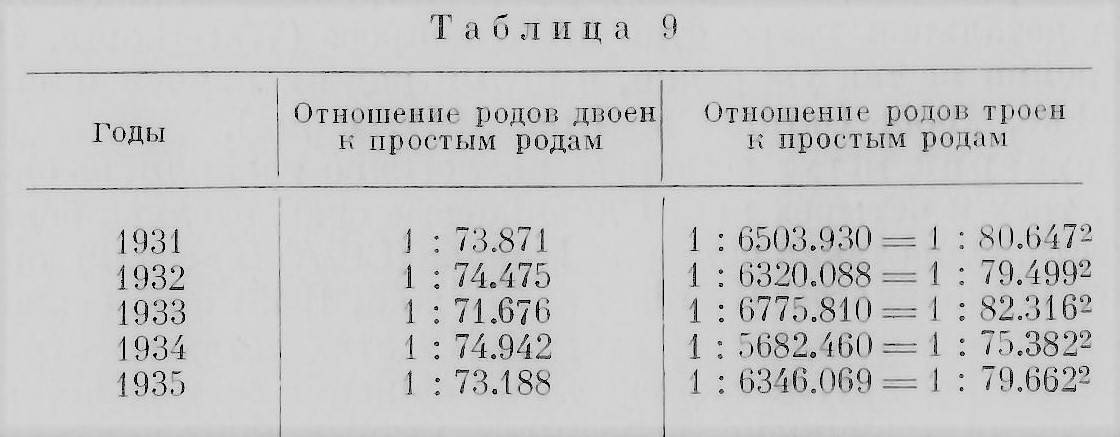 Критикуя закон Эллина, Гедда предлагает назвать его не законом, а принципом. Нет общепринятого объяснения тем отношениям разных типов многоплодия, которые утверждает этот закон или принцип.
Закон Эллина проверялся на статистике рождаемости разных стран. Эти данные, как известно, не соответствуют численности реальных случаев многоплодных беременностей разных типов. Было бы интересно сопоставить числа многоплодных беременностей с числами многоплодных родов и рассмотреть те и другие с точки зрения закона Эллина, который, вероятно, числовые отношения многоплодных зачатий и беременностей не отражает. Получить материал по численности многоплодных беременностей, конечно, трудно, и пока исследования, решающего этот вопрос, нет.
На животных закон Эллина почти не проверялся и подтверждения не нашел (Richter, 1926). Для понимания сущности этого «закона» исследования на животных были бы полезны.
Переходим к рассмотрению частоты рождения более сложных случаев многоплодия — троен, четверен и т. д. По своему составу они могут состоять из разных комбинаций ОБ и РБ, например: тройня может произойти из одного, двух или трех яиц, четверня, соответственно, из одного до четырех яиц, и т. д. Пользуясь законом Эллина, можно попытаться рассчитать возможные количественные отношения в таких случаях (Waterhouse, 1953). Если через «и» обозначить частоту ОБ, а через «v» РБ, то получим:
Критикуя закон Эллина, Гедда предлагает назвать его не законом, а принципом. Нет общепринятого объяснения тем отношениям разных типов многоплодия, которые утверждает этот закон или принцип.
Закон Эллина проверялся на статистике рождаемости разных стран. Эти данные, как известно, не соответствуют численности реальных случаев многоплодных беременностей разных типов. Было бы интересно сопоставить числа многоплодных беременностей с числами многоплодных родов и рассмотреть те и другие с точки зрения закона Эллина, который, вероятно, числовые отношения многоплодных зачатий и беременностей не отражает. Получить материал по численности многоплодных беременностей, конечно, трудно, и пока исследования, решающего этот вопрос, нет.
На животных закон Эллина почти не проверялся и подтверждения не нашел (Richter, 1926). Для понимания сущности этого «закона» исследования на животных были бы полезны.
Переходим к рассмотрению частоты рождения более сложных случаев многоплодия — троен, четверен и т. д. По своему составу они могут состоять из разных комбинаций ОБ и РБ, например: тройня может произойти из одного, двух или трех яиц, четверня, соответственно, из одного до четырех яиц, и т. д. Пользуясь законом Эллина, можно попытаться рассчитать возможные количественные отношения в таких случаях (Waterhouse, 1953). Если через «и» обозначить частоту ОБ, а через «v» РБ, то получим:
р = u + v,тогда
р2 = и2 + 2uv + v2.Через p2, согласно закону Эллина, обозначается общее число троен, u2 представляет частоту однояйцевых троен, 2uv — двуяйцевых, а v2 — трехяйцевых. Эти ожидаемые по расчету отношения, конечно, могут расходиться с наблюдаемыми в связи с различными «случайными» обстоятельствами, как отмирание отдельных компонентов тройни и т. д. Требуется более детальная разработка этого вопроса (Waterhouse, 1953). Тройни не так уж редки, и целый ряд их изучен и описан (Buschke, 1935; Gardner a. Rife, 1941, и др.). Четверни могут состоять, аналогично тройням, из одного, двух, трех и четырех яиц. Однояйцевые сравнительно большая редкость. Одна такая четверня Морлок (США) (рис. 129) описывалась несколько раз (Gardner a. Newman, 1943) и особенно любопытна с психологической стороны. (Об этих детях см. главу одиннадцатую). Описаны и все другие возможные комбинации состава четверни, в частности возникшие из четырех разных яиц, как разного пола, например Гери (Schlaginhaufen, 1940) и Шензе (рис. 130; Gerdner a. Newman, 1944), так и одного пола — мальчики Перрикон (Gardner a. Newman, 1940b). Список всех обследованных и описанных четверен — их всего только 9 — дает Гедда (Gedda, 1951). В этот перечень не входит четверня, описанная врачом Николь в 1935 г. у пигмеев, племени низкорослых негроидов, живущем в лесах экваториальной Африки (Камерун). Из этой четверни выжило 3 девочки, очень похожие друг на друга; им было 3 года 5 месяцев, когда Николь их видел (Caullery, 1945).

 Четверни по полу могут быть четырех мыслимых типов-ММММ, МММЖ, ММЖЖ, МЖЖЖ, ЖЖЖЖ. Была сделана попытка сосчитать, как часто встречается каждый из этих типов (Hamlett, 1935а). Из 48 четверен в США оказалось:
Четверни по полу могут быть четырех мыслимых типов-ММММ, МММЖ, ММЖЖ, МЖЖЖ, ЖЖЖЖ. Была сделана попытка сосчитать, как часто встречается каждый из этих типов (Hamlett, 1935а). Из 48 четверен в США оказалось:
все М — 13 четверен 3М+1Ж — 6 » 2М+2Ж — 12 » 1М+3Ж — 7 » все Ж — 10 »
В общем на 101 женщину приходится 91 мужчина, т. е. 111 : 100, при соотношении полов в населении 106 : 100. Это говорит об уменьшении относительного количества детей мужского пола в многоплодных родах. То же самое отмечает Стокс (Stocks, 1952), объясняя это тенденцией эмбрионов мужского пола к большей смертности при многоплодных беременностях, чем при одиночных. Как уже отмечалось, относительное число мальчиков оказывается меньшим уже при рождении двоен. Эта убыль мальчиков возрастает с числом плодов свыше двух, как видно из табл. 10 (Turpin et Schutzenberger, 1950), приводимой с небольшим сокращением (цифр средних ошибок).
 В табл. 10 выделяется цифра мальчиков в четвернях у французов. Автор объясняет это большим числом четверен мужского пола, однако остается непонятным, чем это вызвано.
Убыль мальчиков подтверждается и данными других авторов (ср. Miettiiien, 1954).
Пятерни, согласно закону Эллина, рождаются, считая n=86, один раз на 54 700 816 родов. За последние 150 лет в США было установлено 4 случая родов пятерни, в Канаде — 2, что в общей соответствует ожидаемым числам. Среди них были и однояйцевые. Всего по литературным данным обнаружена 71 пятерня, из них 56 случаев считается достоверных (Miettinen, 1954).
Поскольку пятерни, по-видимому, рождаются всегда недоносками, они легко погибают, и до недавнего времени не было известно ни одной пятерни, все члены которой прожили хотя бы несколько дней. Пока что в мировой литературе известен один случай пятерни, однояйцевой, все 5 близнецов которой не только выжили в самое трудное для них время первых месяцев жизни, но даже благополучно достигли взрослого возраста. Это знаменитые на весь мир девицы Дионн. На них стоит остановиться и несколько подробнее рассмотреть этот случай. Пятерня эта родилась 28 мая 1934 г. у бедного канадского фермера Дионн, французского происхождения (рис. 131; см. так же рис. 36).
В табл. 10 выделяется цифра мальчиков в четвернях у французов. Автор объясняет это большим числом четверен мужского пола, однако остается непонятным, чем это вызвано.
Убыль мальчиков подтверждается и данными других авторов (ср. Miettiiien, 1954).
Пятерни, согласно закону Эллина, рождаются, считая n=86, один раз на 54 700 816 родов. За последние 150 лет в США было установлено 4 случая родов пятерни, в Канаде — 2, что в общей соответствует ожидаемым числам. Среди них были и однояйцевые. Всего по литературным данным обнаружена 71 пятерня, из них 56 случаев считается достоверных (Miettinen, 1954).
Поскольку пятерни, по-видимому, рождаются всегда недоносками, они легко погибают, и до недавнего времени не было известно ни одной пятерни, все члены которой прожили хотя бы несколько дней. Пока что в мировой литературе известен один случай пятерни, однояйцевой, все 5 близнецов которой не только выжили в самое трудное для них время первых месяцев жизни, но даже благополучно достигли взрослого возраста. Это знаменитые на весь мир девицы Дионн. На них стоит остановиться и несколько подробнее рассмотреть этот случай. Пятерня эта родилась 28 мая 1934 г. у бедного канадского фермера Дионн, французского происхождения (рис. 131; см. так же рис. 36).
 У него уже тогда было 5 человек детей. Не удивительно, что папаша Дионн, увидав, что один за другим стали появляться близнецы, потрясенный, скрылся в лесу. Близнецы родились приблизительно за 2 месяца до срока. Они были очень малы, все впятером весили примерно как один нормальный ребенок. Самая «тяжелая» (Ивонна) весила 2 фунта 14 унций, а самая «легкая» (Мари) — 1 фунт 15 унций.[13] Благодаря инициативе и энергии местного врача Дэфо, а в дальнейшем помощи ряда других лиц новорожденных близнецов удалось спасти. Их появление на свет и ранний период их жизни красочно описал Поль де Крюи в книге «Стоит ли им жить?» (1937). Пятерня Дионн скоро стала знаменита не только в Канаде, но и в США, и в Европе, привлекая массу любопытных, репортеров, ученых и т. д. Благодаря бесчисленным подаркам самого различного рода семья Дионн быстро разбогатела. Мало того, Канаде благодаря туристам пятерня Дионн ежегодно приносила доход около 25 миллионов долларов (Newman, 1940с). Хотя послед пятерни и не сохранился, близнецы являются несомненно однояйцевыми, как это было установлено на основании сходства близнецов по ряду наследственных признаков (McArthur, 1938). Интересно, что между двумя парами в составе пятерни можно установить более тесное сходство, особенно между Мари и Эмилией, которые «зеркально» похожи: одна правша, другая левша (Эмилия, единственная из всех пяти), у одной завиток на темени идет по ходу часовой стрелки, у другой — против хода (Эмилия) и т. д. На основании такого сходства Ньюмен (Newman, 1940с) построил схему (рис. 27) гипотетического образования пятерни Дионн путем повторного раздвоения зиготы по аналогии с образованием близнецов у армадила.
Возможно, что и у Цецилии была также партнерша, погибшая в виде спонтанного аборта, каковой предполагается по некоторым данным на третьем месяце беременности. Пережив первые трудные месяцы, близнецы, поставленные в наилучшие условия, какие только можно было придумать, стали хорошо прибывать в весе. Интересно, что разница в весе между ними постепенно выравнялась (см. главу тринадцатую). Они были похожи между собой по развитию моторики. Речь их отставала: в 3 года они говорили как двухлетние. Отчасти это объясняется тем, что жили они изолированно от внешнего мира и не общались со сверстниками. Между собой они разговаривали на каком-то своеобразном жаргоне, окружение их было однообразно, все их желания предупреждались. При всем наследственном сходстве этих детей и при исключительном сходстве окружающей среды (все пять всегда жили вместе и ни одна из девочек не находилась в привилегированном положении по сравнению с остальными) каждая из пяти приобрела известные индивидуальные особенности, выработала свою собственную личность. Вероятно, это прежде всего связано с тем положением, которое каждая из девочек приобрела в их коллективе пятерни как целого (ср. главу одиннадцатую). К сожалению, этот интересный вопрос мало освещен в литературе, хотя психологией Дионн, как и другими их свойствами, сравнительно много занимались видные ученые (ср. Newman, 1940с). В послевоенные годы о пятерне Дионн писали мало; что они теперь собой представляют, мне неизвестно.
Теперь мы можем вернуться к вопросу о близнецах с большим числом партнеров, чем пять.
По закону Эллина шестерни родятся чрезвычайно редко — примерно один раз на 4712 миллионов родов. Шестерни насчитываются единицами в литературе. Их, по-видимому, установлено около 6 ко времени окончания второй мировой войны (Gates, 1946). К ним присоединился еще один случай, тоже недостаточно достоверный (Newman, 1948). Ни одна шестерня не сохранилась полностью и не была изучена.
Семерни, если можно так выразиться, настолько редки, что о них в литературе почти нет сведений. В Германии (город Гамелн) описан памятник в виде барельефа, воздвигнутый по случаю смерти в 1600 г. семерни, состоявшей из двух мальчиков и пяти девочек и недолго жившей (рис. 132). Все семь младенцев изображены на памятнике (Cook, 1934, и др.). Случай якобы родившейся в 1907 г. семерни описал Томас (Thomas, 1921). По газетам, в Испании в 1943 г. будто бы был случай родов семи живых детей, из которых двое сразу умерли (Caullery, 1945).
У него уже тогда было 5 человек детей. Не удивительно, что папаша Дионн, увидав, что один за другим стали появляться близнецы, потрясенный, скрылся в лесу. Близнецы родились приблизительно за 2 месяца до срока. Они были очень малы, все впятером весили примерно как один нормальный ребенок. Самая «тяжелая» (Ивонна) весила 2 фунта 14 унций, а самая «легкая» (Мари) — 1 фунт 15 унций.[13] Благодаря инициативе и энергии местного врача Дэфо, а в дальнейшем помощи ряда других лиц новорожденных близнецов удалось спасти. Их появление на свет и ранний период их жизни красочно описал Поль де Крюи в книге «Стоит ли им жить?» (1937). Пятерня Дионн скоро стала знаменита не только в Канаде, но и в США, и в Европе, привлекая массу любопытных, репортеров, ученых и т. д. Благодаря бесчисленным подаркам самого различного рода семья Дионн быстро разбогатела. Мало того, Канаде благодаря туристам пятерня Дионн ежегодно приносила доход около 25 миллионов долларов (Newman, 1940с). Хотя послед пятерни и не сохранился, близнецы являются несомненно однояйцевыми, как это было установлено на основании сходства близнецов по ряду наследственных признаков (McArthur, 1938). Интересно, что между двумя парами в составе пятерни можно установить более тесное сходство, особенно между Мари и Эмилией, которые «зеркально» похожи: одна правша, другая левша (Эмилия, единственная из всех пяти), у одной завиток на темени идет по ходу часовой стрелки, у другой — против хода (Эмилия) и т. д. На основании такого сходства Ньюмен (Newman, 1940с) построил схему (рис. 27) гипотетического образования пятерни Дионн путем повторного раздвоения зиготы по аналогии с образованием близнецов у армадила.
Возможно, что и у Цецилии была также партнерша, погибшая в виде спонтанного аборта, каковой предполагается по некоторым данным на третьем месяце беременности. Пережив первые трудные месяцы, близнецы, поставленные в наилучшие условия, какие только можно было придумать, стали хорошо прибывать в весе. Интересно, что разница в весе между ними постепенно выравнялась (см. главу тринадцатую). Они были похожи между собой по развитию моторики. Речь их отставала: в 3 года они говорили как двухлетние. Отчасти это объясняется тем, что жили они изолированно от внешнего мира и не общались со сверстниками. Между собой они разговаривали на каком-то своеобразном жаргоне, окружение их было однообразно, все их желания предупреждались. При всем наследственном сходстве этих детей и при исключительном сходстве окружающей среды (все пять всегда жили вместе и ни одна из девочек не находилась в привилегированном положении по сравнению с остальными) каждая из пяти приобрела известные индивидуальные особенности, выработала свою собственную личность. Вероятно, это прежде всего связано с тем положением, которое каждая из девочек приобрела в их коллективе пятерни как целого (ср. главу одиннадцатую). К сожалению, этот интересный вопрос мало освещен в литературе, хотя психологией Дионн, как и другими их свойствами, сравнительно много занимались видные ученые (ср. Newman, 1940с). В послевоенные годы о пятерне Дионн писали мало; что они теперь собой представляют, мне неизвестно.
Теперь мы можем вернуться к вопросу о близнецах с большим числом партнеров, чем пять.
По закону Эллина шестерни родятся чрезвычайно редко — примерно один раз на 4712 миллионов родов. Шестерни насчитываются единицами в литературе. Их, по-видимому, установлено около 6 ко времени окончания второй мировой войны (Gates, 1946). К ним присоединился еще один случай, тоже недостаточно достоверный (Newman, 1948). Ни одна шестерня не сохранилась полностью и не была изучена.
Семерни, если можно так выразиться, настолько редки, что о них в литературе почти нет сведений. В Германии (город Гамелн) описан памятник в виде барельефа, воздвигнутый по случаю смерти в 1600 г. семерни, состоявшей из двух мальчиков и пяти девочек и недолго жившей (рис. 132). Все семь младенцев изображены на памятнике (Cook, 1934, и др.). Случай якобы родившейся в 1907 г. семерни описал Томас (Thomas, 1921). По газетам, в Испании в 1943 г. будто бы был случай родов семи живых детей, из которых двое сразу умерли (Caullery, 1945).
 Вероятность рождения семерни, а тем более восьмерни, вообще ставится под сомнение. До сих пор, по-видимому, нет еще ни одного случая, достоверно установленного, в печати же появлялись заведомо лживые сообщения такого рода. Примером может служить заметка в одном американском журнале в 1872 г. Об этом случае недавно писал Ньюмен (1948). Оказалось, что это сообщение будто бы злонамеренная фальшивка, написанная отвергнутым поклонником той дамы, которой приписывались столь многоплодные роды (Guttmacker, 1948).
Вероятность рождения семерни, а тем более восьмерни, вообще ставится под сомнение. До сих пор, по-видимому, нет еще ни одного случая, достоверно установленного, в печати же появлялись заведомо лживые сообщения такого рода. Примером может служить заметка в одном американском журнале в 1872 г. Об этом случае недавно писал Ньюмен (1948). Оказалось, что это сообщение будто бы злонамеренная фальшивка, написанная отвергнутым поклонником той дамы, которой приписывались столь многоплодные роды (Guttmacker, 1948).
Глава десятая МОРФОЛОГИЯ И ФИЗИОЛОГИЯ ЧЕЛОВЕЧЕСКИХ БЛИЗНЕЦОВ
Человеческие близнецы, особенно ОБ, изучены во многих отношениях гораздо глубже и разностороннее, чем близнецы животных. Это объясняется не только исключительным интересом к человеку, как объекту изучения взаимодействия факторов наследственности и среды, для чего служат исследования морфологии, физиологии и психологии близнецов в норме и патологии, но и тем удивительным фактом, что относительное количество ОБ у человека оказывается большим, чем у высших млекопитающих, как мы это видели из вышеизложенного материала. Это позволяет изучать не только отдельные «случаи» человеческих близнецов, т. е. единичные пары, как это приходится делать при изучении близнецов у коров, лошадей, обезьян и т. д., но для ряда признаков использовать десятки и даже сотни пар, что дает возможность статистически обрабатывать полученные данные (например, это делалось Коллманом при изучении туберкулеза и шизофрении, см. главу двенадцатую «Заболевания нервной системы»). В таких случаях изучение сходства и различия близнецов требует возможно большего материала. При этом необходимо соблюдать одно очень важное методическое правило, значение которого выяснилось только постепенно, ценой ряда ошибочных выводов. Собирая материал, нельзя ограничиваться только отдельными «интересными» случаями, соответствующими теоретическим предположениям исследователя и потому «нравящимися» ему. Сбор материала должен производиться без отбора в этом смысле, серия пар должна быть «безотборная», даже включать тех близнецов, один из партнеров которых умер (Allen, 1955b). Только в такой безотборной серии изучаемый признак может обнаружиться со свойственным ему многообразием проявления и в тех количественных отношениях, которые соответствуют реальной действительности и поэтому пригодны для суждения о роли наследственности в его проявлении. Так, например, интересуясь наследственностью рака, некоторые исследователи отбирали только те случаи ОБ, когда оба близнеца имели раковую опухоль. Сравнивая такую серию ОБ с серией РБ, где встречаются пары, имеющие только одного ракового больного, делали вывод, что среди ОБ больше конкордантных пар (т. е. внутрипарно похожих по данному признаку), чем среди РБ, а следовательно, ввиду одинаковой наследственности ОБ рак возникает на базе «наследственного предрасположения» к нему и т. д. Сравнение безотборной серии ОБ, т. е. серии, в которую включались все те пары близнецов, где хоть один партнер болел раком, с безотборной серией РБ показало всю ошибочность такого вывода, а вместе с тем и способа сбора материала, к нему приводящего (см. главу двенадцатую «Рак»). Одним из признаков такой безотборности сравнительно большого материала служит количественное отношение пар близнецов в серии ОБ и серии РБ: оно должно приближаться к отношению чисел этих двух типов близнецов в популяции, т. е. число пар РБ должно быть раза в 3—4 больше, чем число пар ОБ. Ряд признаков на близнецах еще не изучен на больших безотборных сериях. Приходится обращаться к работам, сделанным на малом материале, иногда в той или иной степени отобранном, что, конечно, снижает значение статистической обработки такого материала и выводы из него. Это приходится помнить при рассмотрении соответствующих работ и видеть их относительную ценность. Теперь, переходя к конкретному материалу, мы сначала обратимся к некоторым общим признакам, как например рост и вес, легко измеримым, а затем перейдем к рассмотрению отдельных частей тела и органов близнецов. Признаки измеримые, выражающие размеры тела и соотношения частей его, не раз изучались на близнецах различных стран (Verschuer, 1927 — 1954; Бунак, 1926; Dahlberg, 1926; Stocks, 1930; Newman et al., 1937, и др.). В качестве примера такого рода материала рассмотрим таблицу Бунака (1926), составленную на основании обмера московских близнецов в возрасте 13—16 лет при условии, что каждая пара жила приблизительно в одинаковых условиях (табл. 11). Мы видим, что величина М для РБ значительно больше, чем для ОБ, что говорит о большем внутрипарном сходстве ОБ.
Аналогичная картина получена для московских детей в возрасте 8—10 лет (Соболева и Игнатьев, 1936) с помощью другого приема — вычисления коэффициента корреляции между близнецами каждой пары (табл. 12). Как известно, чем больше коэффициент корреляции приближается к единице, тем больше сходство для этих признаков.
В ряде работ внутрипарная разница в общей форме вычисляется с помощью простого приема, предложенного Фершюром (Verschuer, 1927), так называемого процентного отклонения («Е»), Эта величина вдвое меньше, чем вычисленная Бунаком в табл. 11 («М»), так как Фершюр, как мы сейчас увидим из примера, берет половину внутрипарной разницы в процентах к средней величине пары близнецов, а Бунак —не половину разницы, а всю. Для получения процентного уклонения по Фершюру сначала вычисляется средняя величина двух каких-нибудь измерений данной пары близнецов, а затем отклонение от нее в процентах. Например, если один из близнецов имеет рост 168.4 см, а другой 170.2 см, то средняя величина роста обоих будет 169.3 см, а отклонение от нее каждого из близнецов будет равняться 0.9 см. Процентное уклонение данной пары от средней величины их роста будет равно 0.9x100 : 169.3 = 0.53%. Чтобы получить среднее процентное отклонение для нескольких пар близнецов, берется средняя величина процентных отклонений этих пар. Фершюр получил цифры такого среднего процентного отклонения для нескольких признаков обоих типов близнецов из немецкого населения (табл. 13).
Мы видим, что величина М для РБ значительно больше, чем для ОБ, что говорит о большем внутрипарном сходстве ОБ.
Аналогичная картина получена для московских детей в возрасте 8—10 лет (Соболева и Игнатьев, 1936) с помощью другого приема — вычисления коэффициента корреляции между близнецами каждой пары (табл. 12). Как известно, чем больше коэффициент корреляции приближается к единице, тем больше сходство для этих признаков.
В ряде работ внутрипарная разница в общей форме вычисляется с помощью простого приема, предложенного Фершюром (Verschuer, 1927), так называемого процентного отклонения («Е»), Эта величина вдвое меньше, чем вычисленная Бунаком в табл. 11 («М»), так как Фершюр, как мы сейчас увидим из примера, берет половину внутрипарной разницы в процентах к средней величине пары близнецов, а Бунак —не половину разницы, а всю. Для получения процентного уклонения по Фершюру сначала вычисляется средняя величина двух каких-нибудь измерений данной пары близнецов, а затем отклонение от нее в процентах. Например, если один из близнецов имеет рост 168.4 см, а другой 170.2 см, то средняя величина роста обоих будет 169.3 см, а отклонение от нее каждого из близнецов будет равняться 0.9 см. Процентное уклонение данной пары от средней величины их роста будет равно 0.9x100 : 169.3 = 0.53%. Чтобы получить среднее процентное отклонение для нескольких пар близнецов, берется средняя величина процентных отклонений этих пар. Фершюр получил цифры такого среднего процентного отклонения для нескольких признаков обоих типов близнецов из немецкого населения (табл. 13).
 Эти цифры также показывают большее впутрипарное сходство ОБ по сравнению с РБ, а среди РБ — относительно большее сходство близнецов одинакового пола, чем разного.
Среди приведенных признаков одни оказываются более внутрипарно похожими, чем другие, например рост в среднем больше похож, чем вес, и это независимо от типа близнецов. Связь этих признаков с возрастом будет рассмотрена в другом месте (см. глава тринадцатая).
Внутрипарные различия обоих типов близнецов можно для разных признаков изобразить также графически, если, например, на абсциссе отложить числа, характеризующие эти различия, а на ординате — частоты в виде числа пар в процентах, как это сделал Ньюмен с сотрудниками (Newmen et al., 1937) для 50 пар ОБ и 50 пар РБ. В качестве примера мы приводим кривые для роста, длины головы и веса (рис. 133).
Эти цифры также показывают большее впутрипарное сходство ОБ по сравнению с РБ, а среди РБ — относительно большее сходство близнецов одинакового пола, чем разного.
Среди приведенных признаков одни оказываются более внутрипарно похожими, чем другие, например рост в среднем больше похож, чем вес, и это независимо от типа близнецов. Связь этих признаков с возрастом будет рассмотрена в другом месте (см. глава тринадцатая).
Внутрипарные различия обоих типов близнецов можно для разных признаков изобразить также графически, если, например, на абсциссе отложить числа, характеризующие эти различия, а на ординате — частоты в виде числа пар в процентах, как это сделал Ньюмен с сотрудниками (Newmen et al., 1937) для 50 пар ОБ и 50 пар РБ. В качестве примера мы приводим кривые для роста, длины головы и веса (рис. 133).
 Переходим теперь к признакам более частным, зачастую не поддающимся достаточно полному измерению. Сюда относятся различные части лица, уши, особенности кожи и ее производных и другие признаки.
Переходим теперь к признакам более частным, зачастую не поддающимся достаточно полному измерению. Сюда относятся различные части лица, уши, особенности кожи и ее производных и другие признаки.
Морфология головы и рук
Начнем с лица, имеющего в общем важное значение для суждения о сходстве близнецов. Надо прежде всего отметить асимметрию лица (рис. 57), иногда очень заметно выступающую (ср.: Becher, 1935, и др.). Она может быть у близнецов различной: зеркальной или же гомологичной. Кроме промеров лица, о которых уже упоминалось, значение имеют различные детали морфологии лица, не поддающиеся точной оценке с помощью числа. Сюда относятся форма лба, щек, подбородка, общего овала лица и т. п., которые обычно могут быть только описаны и показаны с помощью фотографии. Поэтому они сравнительно хуже изучены у близнецов, хотя на эти признаки всегда обращается внимание. Лучше и точнее удается изучить такие важные части лица, как нос, глаза, рот. К ним мы и переходим.
Ряд морфологических деталей носа можно измерять — высоту, основание, ноздри и т. д., однако полностью его форму числами характеризовать все же нельзя. Внутрипарно ОБ по ряду особенностей формы носа обычно более похожи, чем РБ (Leicher, 1929; Verschuer, 1931/32; Abel, 1932; Gedda, 1951, и др.). Так, например, из 31 пары ОБ лишь у 8 пар найдены небольшие различия формы носа, тогда как у 55% РБ констатирована дискордантность морфологии носа (Leicher, 1928). На ряде фотографий книги Лейхера можно легко убедиться в сходстве характерных особенностей носа у ОБ (ср. рис. 1,160, А и др.), даже не прибегая к специальным измерениям. Внутреннее строение носа близнецов изучалось несколькими исследователями, обнаружившими заметное впутрипарное сходство ОБ (Davenport, 1939; Luchsinger, 1940, 1944, и др.).
 Разрез и размеры рта, толщина и форма губ, высота слизистой оболочки верхней и нижней губы и т. д. — все это признаки, также многократно изучавшиеся у близнецов и имеющие значительно большее внутрипарное сходство у ОБ, чем у РБ (ср.: Lotze, 1937; Gedda, 1951, и др.). О зубах речь будет ниже.
Цвет радужной оболочки глаз (ириса) у пары ОБ, как правило, очень похож, часто неразличим. И не только цвет, но и характер распределения пигмента радужной разных оттенков, то, что можно условно назвать «рисунком» или «структурой» ее, также обычно внутрипарно чрезвычайно похож (рис. 134). Поскольку здесь речь идет о цветовом феномене, простая фотография лишь очень приблизительно передает его, в действительности же это сходство гораздо заметнее. Оно настолько постоянно и точно, что служит одним из критериев диагноза однояйцевости близнецов (Schwaegerle, 1938; Essen-Мollег, 1941; Gedda, 1951, и др.). И цвет и «структура» радужной могут быть определены по специальным шкалам и схемам сравнительно точно, хотя и без количественного анализа (Бунак, 1941).
Разрез и размеры рта, толщина и форма губ, высота слизистой оболочки верхней и нижней губы и т. д. — все это признаки, также многократно изучавшиеся у близнецов и имеющие значительно большее внутрипарное сходство у ОБ, чем у РБ (ср.: Lotze, 1937; Gedda, 1951, и др.). О зубах речь будет ниже.
Цвет радужной оболочки глаз (ириса) у пары ОБ, как правило, очень похож, часто неразличим. И не только цвет, но и характер распределения пигмента радужной разных оттенков, то, что можно условно назвать «рисунком» или «структурой» ее, также обычно внутрипарно чрезвычайно похож (рис. 134). Поскольку здесь речь идет о цветовом феномене, простая фотография лишь очень приблизительно передает его, в действительности же это сходство гораздо заметнее. Оно настолько постоянно и точно, что служит одним из критериев диагноза однояйцевости близнецов (Schwaegerle, 1938; Essen-Мollег, 1941; Gedda, 1951, и др.). И цвет и «структура» радужной могут быть определены по специальным шкалам и схемам сравнительно точно, хотя и без количественного анализа (Бунак, 1941).
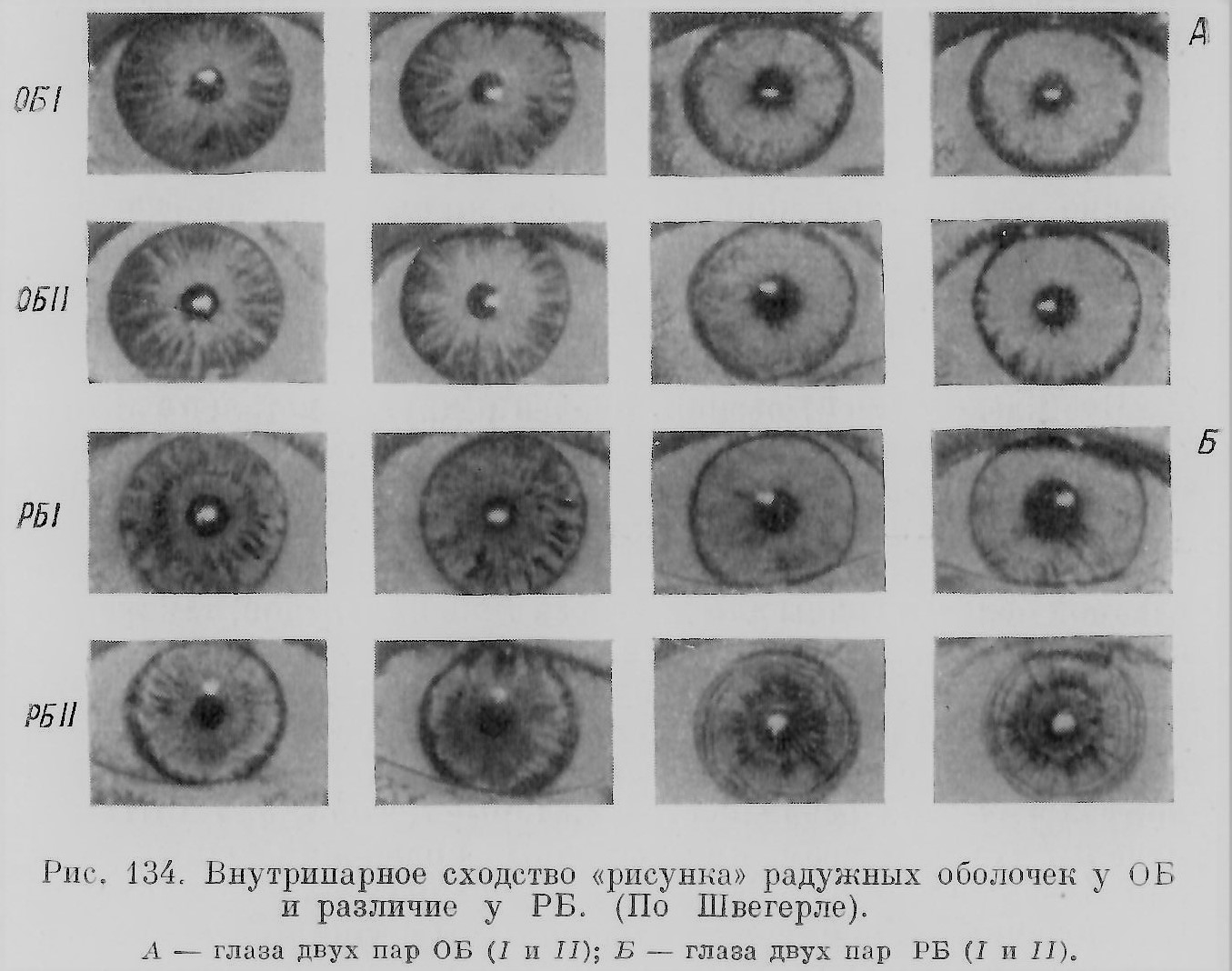 Известно, что пигментация радужной оболочки правого и левого глаза того же субъекта хотя и редко, но различается; обычно это различие временного характера. Приблизительно такого же порядка различия могут существовать между глазами пары ОБ. Так, из 256 пар ОБ полное сходство пигментации ириса оказалось у 86.7%, небольшие различия — у 12.9%, и лишь у одной пары (0.4%) было заметное различие цвета, которое впоследствии исчезло. Наоборот, среди РБ только у 13% установлено полное сходство, у 18% небольшое различие, а у 72% цвет радужной был заметно разный (Lotze, 1937). Аналогичные данные установлены и на другом европейском материале (Dahlberg, 1926; Waardenburg, 1932, 1950, и др.).
Возрастная изменчивость цвета глаз у близнецов еще мало изучена. На почве такой изменчивости, вероятно, могут возникать некоторые внутрипарные различия пигментации радужной У ОБ.
Изучались также некоторые детали анатомии глазного яблока близнецов. Оказалось, что особенности строения края зрачка имеют большое внутрипарное сходство у ОБ. Глазное дно также обнаруживает подобное сходство и т. д. (Huber, 1931; Jancke, 1941; Lehrfield, 1948, и др.). Ряд патологических процессов в глазу, в частности связанных со старостью, протекают внутрипарно похоже у ОБ (A. Vogt et al., 1939, и др.).
Косоглазие, сравнительно мало еще изученное у близнецов, обычно встречается конкордантно у пары ОБ, иногда зеркально, т. е. у одного близнеца косит левый глаз, а у другого — правый (Steinmann, 1943, и др.). У РБ, наоборот, преобладает внутрипарная дискордантность, т. е. один косит, а другой не косит.
Что касается век, ресниц, бровей и других деталей глазной области, то относительно этих признаков установлено заметно большее внутрипарное сходство ОБ по сравнению с РБ (Sieder, 1938; Jancke, 1941; Gedda, 1951, и др.).
Некоторые исследователи считают, что эти мелкие признаки глазной области годны для диагноза типа близнецов, так же как и особенности глазного яблока.
О функциях глаза речь будет в другом месте (см. далее «Физиология анализаторов»).
Уши, наконец, также являются очень важным признаком для суждения о сходстве близнецов. Наружное ухо имеет ряд морфологических особенностей, которые легко могут быть измерены, что дает возможность точных сравнений ушных раковин близнецов. Можно сравнивать оба уха одного субъекта, гомологичные уши близнецов одной пары и уши противоположных сторон (правое первого близнеца с левым второго и левое первого с правым второго). Для сравнения удобнее брать не абсолютные цифры, а среднюю разность, как это нередко делают. Не входя в детали таких сравнений, рассмотрим лишь несколько итоговых цифр, полученных путем обобщения средней разности 19 различных промеров (в мм) ушной раковины (Quelprud, 1932) и сведенных в табл. 14.
Известно, что пигментация радужной оболочки правого и левого глаза того же субъекта хотя и редко, но различается; обычно это различие временного характера. Приблизительно такого же порядка различия могут существовать между глазами пары ОБ. Так, из 256 пар ОБ полное сходство пигментации ириса оказалось у 86.7%, небольшие различия — у 12.9%, и лишь у одной пары (0.4%) было заметное различие цвета, которое впоследствии исчезло. Наоборот, среди РБ только у 13% установлено полное сходство, у 18% небольшое различие, а у 72% цвет радужной был заметно разный (Lotze, 1937). Аналогичные данные установлены и на другом европейском материале (Dahlberg, 1926; Waardenburg, 1932, 1950, и др.).
Возрастная изменчивость цвета глаз у близнецов еще мало изучена. На почве такой изменчивости, вероятно, могут возникать некоторые внутрипарные различия пигментации радужной У ОБ.
Изучались также некоторые детали анатомии глазного яблока близнецов. Оказалось, что особенности строения края зрачка имеют большое внутрипарное сходство у ОБ. Глазное дно также обнаруживает подобное сходство и т. д. (Huber, 1931; Jancke, 1941; Lehrfield, 1948, и др.). Ряд патологических процессов в глазу, в частности связанных со старостью, протекают внутрипарно похоже у ОБ (A. Vogt et al., 1939, и др.).
Косоглазие, сравнительно мало еще изученное у близнецов, обычно встречается конкордантно у пары ОБ, иногда зеркально, т. е. у одного близнеца косит левый глаз, а у другого — правый (Steinmann, 1943, и др.). У РБ, наоборот, преобладает внутрипарная дискордантность, т. е. один косит, а другой не косит.
Что касается век, ресниц, бровей и других деталей глазной области, то относительно этих признаков установлено заметно большее внутрипарное сходство ОБ по сравнению с РБ (Sieder, 1938; Jancke, 1941; Gedda, 1951, и др.).
Некоторые исследователи считают, что эти мелкие признаки глазной области годны для диагноза типа близнецов, так же как и особенности глазного яблока.
О функциях глаза речь будет в другом месте (см. далее «Физиология анализаторов»).
Уши, наконец, также являются очень важным признаком для суждения о сходстве близнецов. Наружное ухо имеет ряд морфологических особенностей, которые легко могут быть измерены, что дает возможность точных сравнений ушных раковин близнецов. Можно сравнивать оба уха одного субъекта, гомологичные уши близнецов одной пары и уши противоположных сторон (правое первого близнеца с левым второго и левое первого с правым второго). Для сравнения удобнее брать не абсолютные цифры, а среднюю разность, как это нередко делают. Не входя в детали таких сравнений, рассмотрим лишь несколько итоговых цифр, полученных путем обобщения средней разности 19 различных промеров (в мм) ушной раковины (Quelprud, 1932) и сведенных в табл. 14.

 Внутрипарное сходство ОБ по промерам ушной раковины в общем такое же, как ушей одного субъекта, тогда как у РБ разница заметно больше. Сходство ушных раковин ОБ по сравнению с РБ заметно на глаз, даже без измерений (рис. 135).
Можно также измерить угол, под которым расположена ушная раковина к голове, и в этом отношении внутрипарное сходство ОБ приближается к сходству между ушами одного субъекта и превышает сходство РБ.
Специально изучался Дарвиновский бугорок на ушной раковине (Verschuer, 1931; Quelprud, 1934), ямка у верхнего края наружного уха (Quelprud, 1940) и другие мелкие особенности морфологии ушной раковины близнецов.
Сходство близнецов в отношении деталей строения среднего и внутреннего уха еще мало изучено (Leicher, 1929; Luscher, 1944, и др.). Интересно отметить, что микроскопическое строение барабанной перепонки по ряду деталей имеет заметно большее внутрипарное сходство у ОБ, чем у РБ (Luscher, 1944).
Физиология слуха у близнецов еще почти не исследована (см. далее «Физиология анализаторов»).
Из других частей тела после лица очень существенную роль имеют для сравнения руки, именно кисти рук, на выразительность которых давно уже обращали внимание художники, например Леонардо да Винчи. Здесь важны особенности формы пальцев, ногтей, кожного рельефа дистальных фаланг и ладоней. Последнее имеет очень большое значение при диагнозе близнецов.
Внутрипарное сходство ОБ по промерам ушной раковины в общем такое же, как ушей одного субъекта, тогда как у РБ разница заметно больше. Сходство ушных раковин ОБ по сравнению с РБ заметно на глаз, даже без измерений (рис. 135).
Можно также измерить угол, под которым расположена ушная раковина к голове, и в этом отношении внутрипарное сходство ОБ приближается к сходству между ушами одного субъекта и превышает сходство РБ.
Специально изучался Дарвиновский бугорок на ушной раковине (Verschuer, 1931; Quelprud, 1934), ямка у верхнего края наружного уха (Quelprud, 1940) и другие мелкие особенности морфологии ушной раковины близнецов.
Сходство близнецов в отношении деталей строения среднего и внутреннего уха еще мало изучено (Leicher, 1929; Luscher, 1944, и др.). Интересно отметить, что микроскопическое строение барабанной перепонки по ряду деталей имеет заметно большее внутрипарное сходство у ОБ, чем у РБ (Luscher, 1944).
Физиология слуха у близнецов еще почти не исследована (см. далее «Физиология анализаторов»).
Из других частей тела после лица очень существенную роль имеют для сравнения руки, именно кисти рук, на выразительность которых давно уже обращали внимание художники, например Леонардо да Винчи. Здесь важны особенности формы пальцев, ногтей, кожного рельефа дистальных фаланг и ладоней. Последнее имеет очень большое значение при диагнозе близнецов.
Кожа
Кожа и ее производные — волосы, ногти и проч. — являются легко наблюдаемыми особенностями человеческого организма, имеющими очень важное значение для различения типов близнецов. Прежде всего обратимся к вопросу о цвете кожи. У европейских народов имеется множество оттенков цвета кожи, также у некоторых азиатских. У народностей монголоидной и негроидной расы различий оттенков цвета кожи меньше, в ряде случаев такие различия вовсе отсутствуют. Поэтому у африканских негров, например, невозможно найти какие-либо различия цвета кожи не только между близнецами, но, вероятно, людьми целого племени. Иная картина наблюдается для большинства народностей Европы. ОБ европеоидов обычно имеют очень заметное внутрипарное сходство цвета кожи; зачастую это сходство очень велико, при отсутствии, конечно, каких-нибудь специальных воздействий на одного из близнецов, например солнечного света, вызывающего загар у одного, и тому подобных случаев. Внутрипарная разница у РБ по цвету кожи, наоборот, наблюдается сравнительно часто, например среди немецкого населения более чем у 50% пар РБ (Lotze, 1937). Среди особенностей кожи надо отметить веснушки. У ОБ не найдено дискордантных пар, т. е. если веснушки есть у одного из близнецов, то они есть и у другого, но могут быть, в зависимости от некоторых условий, выражены слабее или сильнее. Так, например, это очевидно из следующих цифр (Verschuer, 1931/32).ОБ.......54++ 16+(+) 0+- 39-- РБ.......10++ 27+(+) 29+- 41--Здесь ++ значит, что веснушки есть у обоих близнецов и одинаково сильно выраженные, +(+), что они есть у обоих, но у одного выражены слабее, +- значит, что у одного веснушек нет, -- что их нет у обоих. Родинки имеют меньшую степень конкордантности и далеко не всегда обнаруживаются на гомологичных местах у ОБ или в одинаковом числе, и т. д. Все же внутрипарное сходство ОБ в этом отношении больше, чем РБ. Коэффициент внутрипарной корреляции для ОБ +0.78, а для РБ +0.31 (Verschuer, 1931/32). Ряд других мелких особенностей и аномалий кожи изучался Сименсом (Siemens, 1924) и другими (см. Gedda, 1951). Волосы, прежде всего головы, легко различаются по цвету и форме (прямые, волнистые, курчавые). У ОБ, как правило, и цвет и форма волос весьма похожи. Однако по оттенку окраски некоторые внутрипарные различия ОБ иногда наблюдаются; например, среди немецкого населения до 10% пар ОБ имеют внутрипарное различие по оттенку цвета волос, тогда как среди РБ того же населения установлено свыше 70% внутрипарного несходства не только по оттенку, но и по цвету. Возрастная изменчивость цвета волос ОБ еще мало изучена. Констатированы редкие случаи заметной внутрипарной разницы по цвету волос у ОБ (Loewy, 1925, и др.). Остается невыясненным, насколько такие различия ОБ оказываются в пределах возрастных изменений или даже индивидуальных, поскольку у одного и того же субъекта одновременно на разных участках головы оттенки волос могут быть заметно различны. Не ясно также, чем бывают вызваны наблюдавшиеся внутрипарные различия цвета волос у ОБ. Но это в общем явления редкие, порядка исключений из правила большого сходства ОБ по цвету волос. По форме волосы у ОБ, как правило, не имеют внутрипарной разницы, тогда как она у РБ есть, хотя встречается реже, чем разница по цвету волос. Так, среди европейского населения преобладает гладкая форма волос, среди негров — курчавая, а потому понятно, что и пара ОБ и пара РБ будет иметь ту же форму волос. Интересно отметить, что волосы головы на темени расположены в виде завитка, который у большинства людей направлен по ходу часовой стрелки. Реже он направлен в обратную сторону, и еще реже встречается двойной завиток или другие особенности расположения волос. Процент дискордантных пар ОБ по этому признаку оказался несколько больше, чем пар РБ, что дало Ньюмену повод считать, что в этой «зеркальности» проявляется первичная двусторонняя симметрия исходного эмбриона, из которого возникли данные ОБ. Однако такая дискордантность ОБ статистически недостаточно обоснована, что побудило Фершюра (Verschuer, 1932) возражать против гипотезы Ньюмена. Требуется дальнейшее исследование этого вопроса. Волосяной покров различных частей головы и тела близнецов также изучался. Например, установлено три типа направления мелких волосков, растущих на лбу, особенно у детей: вниз, вверх и комбинированных из двух первых направлений. Изучение 30 пар ОБ из норвежского населения показало, что у них имеется 100% конкордантности по этому признаку, тогда как у 28 пар РБ найдено только 43% конкордантности (Kiil, 1948). Из других особенностей проявления волосяного покрова на лице существенное значение для диагноза имеют брови и ресницы. Брови могут различаться по ряду свойств: положению, изгибу, ширине, цвету, расположению волос — завитки и т. п. (Breitinger, 1955, и др.). Внутрипарное сходство ОБ в отношении бровей заметно больше, чем РБ, среди которых у некоторых групп европейского населения установлено около 50% дискордантных пар. Ресницы различаются по длине, форме, расположению и т. д., и внутрипарное сходство ОБ в этом отношении явно больше, чем РБ (ср. Gedda, 1951). Волосяной покров тела у близнецов изучен хуже, чем волосы головы, и мы на нем останавливаться не будем. Ногти у ОБ обычно очень похожи, по-видимому, больше, чем у РБ. Нет еще достаточно точных и четких приемов описания и характеристики ногтей для суждения о степени их сходства. Довольно много изучались капилляры кожи, в частности ногтевого ложа (Mayer-List u. НuЬеnег, 1925; Бунак, 1936; Granel, 1937; Schiller, 1937; Lehmann u. Hartlieb, 1937, и др.). Изменчивость капилляров кожи сравнительно велика, и у разных индивидов можно констатировать заметные различия в этом отношении. Внутрипарное сходство ОБ по этому признаку выступает очень отчетливо: по разным данным, у них имеется до 84—100% конкордантности, тогда как у РБ наблюдается от 72% и больше дискордантностн (рис. 136).
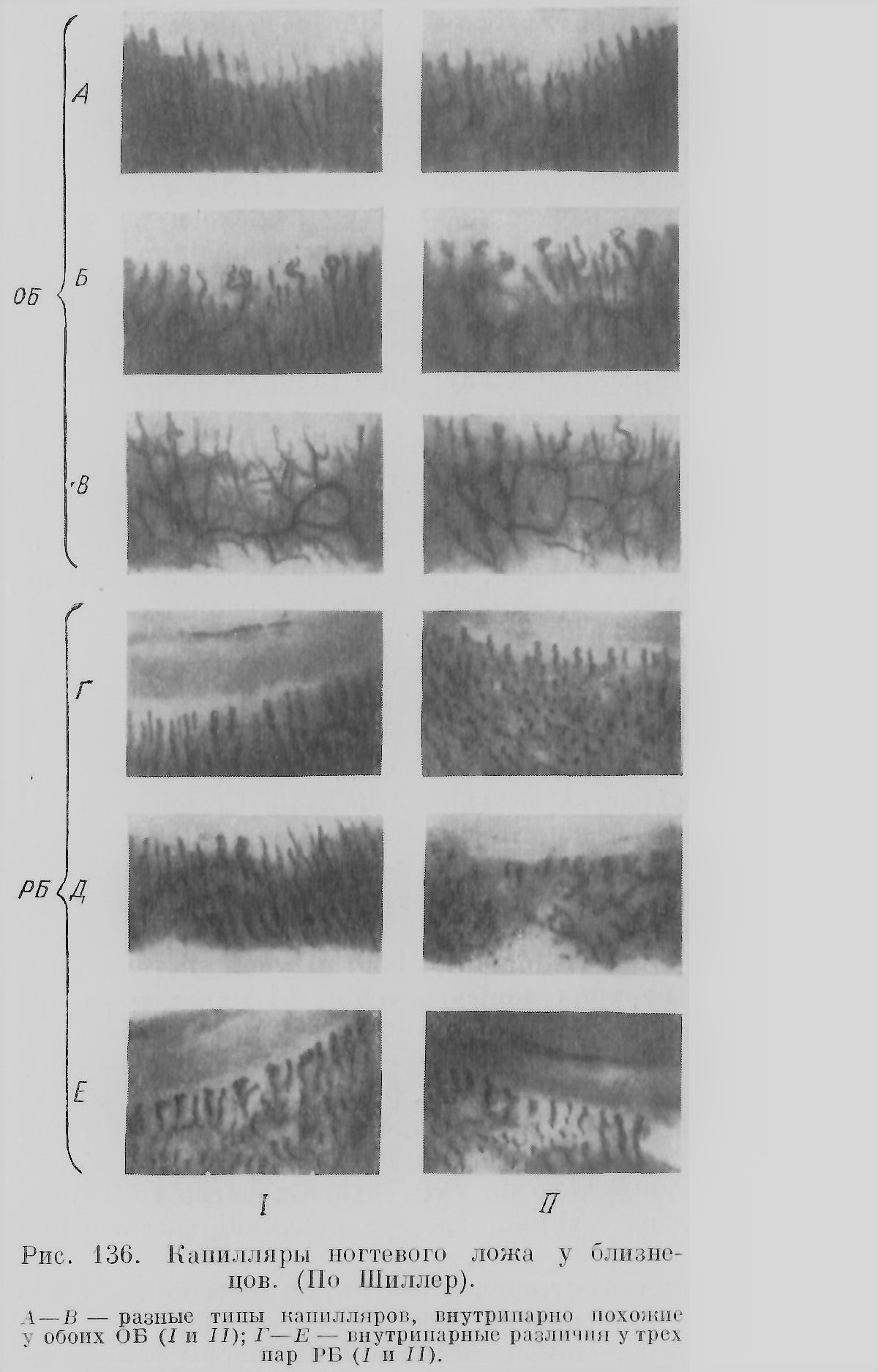 Почти не изучены другие формы кожных сосудов. Имеется лишь одно исследование вен верхних конечностей близнецов-школьников, сделанное Кадановым (Kadanoff, 1939). Вены изучались с помощью наложения манжетки, служащей для измерения кровяного давления. Установлено три основных типа ветвления вен. Оказалось, что у 24 субъектов из 60 исследованных, т. е. у 40%, кожные вены правой и левой руки были разного типа. Лишь у 28 человек, т. е. у 46.7%, обе руки имели один и тот же тип. У остальных близнецов, т. е. у 8 человек, или 13.3% случаев, одна рука имела смешанный тип вен, а другая — один из трех типов. У субъектов одиночек, не близнецов, тип вен правой и левой руки был разный примерно у 45%.
Из 8 пар ОБ у 6, т. е. 75%, гомологичные руки имели тот же тип вен или частично тот же как для правой, так и для левой руки. Среди РБ это наблюдалось в 27.3% случаев, что свидетельствует о значительно большем числе пар с внутрипарным различием. Вообще же мелкое ветвление вен имеет множество видоизменений индивидуального характера даже при наличии общности типа основных вен двух рук.
На двух парах зародышей ОБ (5—7 месяцев) также описано много внутрипарных вариаций кожных вен рук и других частей тела; более крупные вены и артерии мало изменчивы. Возможно, что дискордантное развитие периферических сосудов конечностей зависит от различия положения конечностей зародыша (Siebert, 1937).
Изучение ветвления вен наружной поверхности кистей рук у близнецов показало, что у одной части пар ОБ очень часто обнаруживается ясная симметрия в ветвлении вен правой и левой руки. У другой части пар ОБ симметрия между правой и левой рукой незаметна, но ясно выступает известное сходство ветвления гомологичных рук, т. е. обнаруживается конкордантная асимметрия. Такая асимметрия наблюдается не только на венах, но и на форме лица тех же ОБ. У РБ никакого внутрипарного сходства ветвления вен не установлено (Freerksen, 1938). Чем объяснить существование ОБ с симметричным и асимметричным ветвлением вен, сказать трудно. Это частный вопрос из области той большой проблемы, о которой речь была выше.
Каданов же (Kadanoff, 1939) изучал ветвление кожных нервов верхних конечностей на 11 парах мертворожденных близнецов, частью доношенных. Принадлежность близнецов к одному из двух типов определялась на основании оболочек. 3 пары были признаны ОБ, 5 пар РБ, у 3 пар, за отсутствием данных об оболочках, диагноз поставить нельзя было. И в отношении ветвления нервов найдены различия между правой и левой рукой того же субъекта. Сходство гомологичных рук пары ОБ, при наличии мелких различий, несомненно больше, чем пары РБ (рис. 137).
Почти не изучены другие формы кожных сосудов. Имеется лишь одно исследование вен верхних конечностей близнецов-школьников, сделанное Кадановым (Kadanoff, 1939). Вены изучались с помощью наложения манжетки, служащей для измерения кровяного давления. Установлено три основных типа ветвления вен. Оказалось, что у 24 субъектов из 60 исследованных, т. е. у 40%, кожные вены правой и левой руки были разного типа. Лишь у 28 человек, т. е. у 46.7%, обе руки имели один и тот же тип. У остальных близнецов, т. е. у 8 человек, или 13.3% случаев, одна рука имела смешанный тип вен, а другая — один из трех типов. У субъектов одиночек, не близнецов, тип вен правой и левой руки был разный примерно у 45%.
Из 8 пар ОБ у 6, т. е. 75%, гомологичные руки имели тот же тип вен или частично тот же как для правой, так и для левой руки. Среди РБ это наблюдалось в 27.3% случаев, что свидетельствует о значительно большем числе пар с внутрипарным различием. Вообще же мелкое ветвление вен имеет множество видоизменений индивидуального характера даже при наличии общности типа основных вен двух рук.
На двух парах зародышей ОБ (5—7 месяцев) также описано много внутрипарных вариаций кожных вен рук и других частей тела; более крупные вены и артерии мало изменчивы. Возможно, что дискордантное развитие периферических сосудов конечностей зависит от различия положения конечностей зародыша (Siebert, 1937).
Изучение ветвления вен наружной поверхности кистей рук у близнецов показало, что у одной части пар ОБ очень часто обнаруживается ясная симметрия в ветвлении вен правой и левой руки. У другой части пар ОБ симметрия между правой и левой рукой незаметна, но ясно выступает известное сходство ветвления гомологичных рук, т. е. обнаруживается конкордантная асимметрия. Такая асимметрия наблюдается не только на венах, но и на форме лица тех же ОБ. У РБ никакого внутрипарного сходства ветвления вен не установлено (Freerksen, 1938). Чем объяснить существование ОБ с симметричным и асимметричным ветвлением вен, сказать трудно. Это частный вопрос из области той большой проблемы, о которой речь была выше.
Каданов же (Kadanoff, 1939) изучал ветвление кожных нервов верхних конечностей на 11 парах мертворожденных близнецов, частью доношенных. Принадлежность близнецов к одному из двух типов определялась на основании оболочек. 3 пары были признаны ОБ, 5 пар РБ, у 3 пар, за отсутствием данных об оболочках, диагноз поставить нельзя было. И в отношении ветвления нервов найдены различия между правой и левой рукой того же субъекта. Сходство гомологичных рук пары ОБ, при наличии мелких различий, несомненно больше, чем пары РБ (рис. 137).
 Материал Каданова невелик, и бесспорно обоснованные обобщения на основании его едва ли можно делать. По впечатлению автора, одноименные кожные нервы гомологичных рук имеют меньшую изменчивость, чем одноименные вены.
Переходим к очень важному для диагноза типа близнецов признаку — кожному рельефу, его «узорам». Поверхность кожи имеет своеобразный рельеф: на дистальных фалангах пальцев рук и ног, на ладонях, стопах и т. д. можно легко обнаружить определенные закономерно расположенные «узоры». Эти узоры образованы гребнями, чередующимися с бороздами. Гребни образованы выступами наружного рогового слоя кожи над аналогичными возвышениями лежащей под ними соединительнотканной части кожи, называемыми сосочками и валиками, а борозды — углублениями между гребнями. Поэтому узоры, образованные гребнями, называют также папиллярными (сосочковыми) узорами. Область антропологии, изучающую кожный рельеф, иногда называют особым термином — дерматоскопия (Волоцкой, 1937), а раздел ее, занимающийся только узорами дистальных фаланг пальцев — дактилоскопией, имеющей, как известно, широкое применение в судебной медицине при идентификации личности преступника. Дело в том, что папиллярные узоры, формирующиеся на третьем месяце развития эмбриона, сохраняются без существенного изменения в течение всей жизни человека, совокупность же особенностей этих узоров всех десяти пальцев человека такова, что по ним можно всегда отличить одного субъекта от другого, и не установлено ни одного случая, когда бы у двух людей все отпечатки пальцев обеих рук полностью совпадали. Это обстоятельство вызывает, конечно, интерес к изучению дактилоскопии близнецов и вообще их дерматоглифики, т. е. кожного рельефа.
Материал Каданова невелик, и бесспорно обоснованные обобщения на основании его едва ли можно делать. По впечатлению автора, одноименные кожные нервы гомологичных рук имеют меньшую изменчивость, чем одноименные вены.
Переходим к очень важному для диагноза типа близнецов признаку — кожному рельефу, его «узорам». Поверхность кожи имеет своеобразный рельеф: на дистальных фалангах пальцев рук и ног, на ладонях, стопах и т. д. можно легко обнаружить определенные закономерно расположенные «узоры». Эти узоры образованы гребнями, чередующимися с бороздами. Гребни образованы выступами наружного рогового слоя кожи над аналогичными возвышениями лежащей под ними соединительнотканной части кожи, называемыми сосочками и валиками, а борозды — углублениями между гребнями. Поэтому узоры, образованные гребнями, называют также папиллярными (сосочковыми) узорами. Область антропологии, изучающую кожный рельеф, иногда называют особым термином — дерматоскопия (Волоцкой, 1937), а раздел ее, занимающийся только узорами дистальных фаланг пальцев — дактилоскопией, имеющей, как известно, широкое применение в судебной медицине при идентификации личности преступника. Дело в том, что папиллярные узоры, формирующиеся на третьем месяце развития эмбриона, сохраняются без существенного изменения в течение всей жизни человека, совокупность же особенностей этих узоров всех десяти пальцев человека такова, что по ним можно всегда отличить одного субъекта от другого, и не установлено ни одного случая, когда бы у двух людей все отпечатки пальцев обеих рук полностью совпадали. Это обстоятельство вызывает, конечно, интерес к изучению дактилоскопии близнецов и вообще их дерматоглифики, т. е. кожного рельефа.
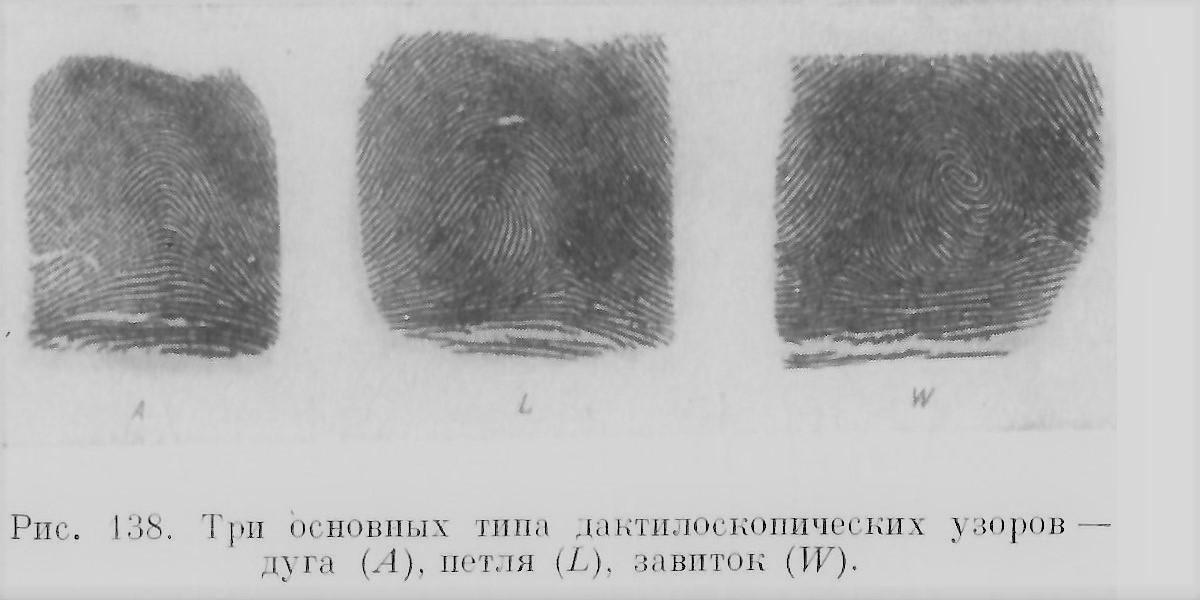 Папиллярные узоры различают по двум особенностям: по типу узора и по числу линий, которыми образован узор. Согласно классификации, установленной еще Гальтоном (Galton, 1895), все многообразие форм узоров сводится к трем основным типам: дуга (А), петля (L) и круг (завиток) (W). Эти типы обычно обозначаются начальной буквой их английского названия, приведенной в скобках (рис. 138). В дуге линии идут поперек дистальной фаланги (с ладонной поверхности), более или менее поднимаясь дистально по середине фаланги. В петле линни в середине узора поворачиваются и следуют назад. Большинство петель открыто в ульнарную сторону, т. е. к мизинцу, и потому обозначаются буквой U, а меньшинство в обратную, радиальную сторону, т. е. к большому пальцу, и обозначаются через R. В петлях есть место встречи трех ходов линий, обозначаемое «дельтой» (напоминает по форме греческую букву Д). Обычно от дельты до центра петли по прямой отсчитывают количество полосок — это числовой (количественный) показатель узора. Так как дуга не имеет дельты, то этот показатель ее считается обычно равным нулю. Наконец, третий тип — круг или завиток — имеет обычно две дельты. Количественный показатель завитка часто берется только по дельте, более удаленной от центра фигуры. Таким образом, три типа дактилоскопических узоров легко различимы и по числу дельт: у дуги их нет, у петли одна, у завитка две дельты. Так эти три типа различал еще отец дактилоскопии Е. Пуркинье (см. Гутскунст, 1955).
Кроме этих трех основных типов, существуют еще некоторые переходные или измененные формы их, сравнительно редко встречающиеся, как например двойной завиток и другие. На них нет надобности останавливаться здесь подробнее.
Детально изучены также узоры на ладонях и ступнях (рис. 139, 140), им также даются условные числовые обозначения, с помощью которых их можно сравнивать (подробности см.: Бунак, 1941; Newman, 1931; Meyer-Heydenhagen, 1934; Geipel, 1942; Schiller, 1942: Cummins a. Midlo, 1943, и др.).
Папиллярные узоры различают по двум особенностям: по типу узора и по числу линий, которыми образован узор. Согласно классификации, установленной еще Гальтоном (Galton, 1895), все многообразие форм узоров сводится к трем основным типам: дуга (А), петля (L) и круг (завиток) (W). Эти типы обычно обозначаются начальной буквой их английского названия, приведенной в скобках (рис. 138). В дуге линии идут поперек дистальной фаланги (с ладонной поверхности), более или менее поднимаясь дистально по середине фаланги. В петле линни в середине узора поворачиваются и следуют назад. Большинство петель открыто в ульнарную сторону, т. е. к мизинцу, и потому обозначаются буквой U, а меньшинство в обратную, радиальную сторону, т. е. к большому пальцу, и обозначаются через R. В петлях есть место встречи трех ходов линий, обозначаемое «дельтой» (напоминает по форме греческую букву Д). Обычно от дельты до центра петли по прямой отсчитывают количество полосок — это числовой (количественный) показатель узора. Так как дуга не имеет дельты, то этот показатель ее считается обычно равным нулю. Наконец, третий тип — круг или завиток — имеет обычно две дельты. Количественный показатель завитка часто берется только по дельте, более удаленной от центра фигуры. Таким образом, три типа дактилоскопических узоров легко различимы и по числу дельт: у дуги их нет, у петли одна, у завитка две дельты. Так эти три типа различал еще отец дактилоскопии Е. Пуркинье (см. Гутскунст, 1955).
Кроме этих трех основных типов, существуют еще некоторые переходные или измененные формы их, сравнительно редко встречающиеся, как например двойной завиток и другие. На них нет надобности останавливаться здесь подробнее.
Детально изучены также узоры на ладонях и ступнях (рис. 139, 140), им также даются условные числовые обозначения, с помощью которых их можно сравнивать (подробности см.: Бунак, 1941; Newman, 1931; Meyer-Heydenhagen, 1934; Geipel, 1942; Schiller, 1942: Cummins a. Midlo, 1943, и др.).

 Многие ученые занимались дерматоглификой близнецов и спорили о степени важности показателей рельефа кожи для диагностики близнецов. Некоторые из специалистов, например Камминс (Cummins, 1930) и Райф (Rife, 1933b; Rife a. Cummins, 1943), считали их ненадежными признаками для этой цели. Их противники, наоборот, стремились доказать существенное значение этих признаков (Komai, 1928; Newman, 1930; Stocks,. 1930, 1933; Verschuer, 1933; Волоцкой, 1936; McArthur, 1938; Slater, 1953; Herrman, 1953; Wendt, 1955, и др.). В результате дактилоскопия близнецов оказалась сравнительно хорошо изученной на значительном материале в несколько сот пар близнецов и достигнуты некоторые интересные обобщения, которые говорят за то, что рельеф кожи несомненно годится для суждения о природе близнецов наравне с другими существенными для диагноза признаками. Одно из важнейших обобщений заключается в следующем «правиле», установленном Комаи и Ньюменом: в большинстве случаев отпечатки и пальцев и ладоней гомологичных рук двух однояйцевых близнецов более похожи по своим рисункам, чем правой и левой руки того же субъекта (рис. 139). Обратное, в общем, можно сказать относительно РБ (Komai, 1928; Newman, 1930, и др.).
Если подсчитать количество полосок дактилоскопических узоров и вычислить коэффициенты корреляции между правой и левой рукой какого-нибудь субъекта, взятого как из числа ОБ, так и РБ (Newman, 1930, и др.), то получится в среднем одинаковое число (r = +0.93(+/-)0.01), свидетельствующее о большой корреляции. Такова же корреляция между левыми руками ОБ одной пары и несколько меньше (r = +0.92(+/-)0.01) между правыми руками. Больше всего коэффициент корреляции (r = +0.95(+/-)0.01) между обеими руками одного из близнецов с обеими руками другого. Для РБ этот коэффициент корреляции значительно меньше (r = +0.46(+/-)0.08).
Однако было бы ошибкой думать, что внутрипарное сходство кожных узоров ОБ всегда одинаково велико и что у них всегда гомологичные пальцы имеют тождественный узор и тот же числовой показатель. Полного тождества никогда не наблюдается, и всегда можно найти известные, хотя бы небольшие, различия между узорами гомологичных пальцев, даже в случае исключительно большого сходства, как например на отпечатках рис. 141.
Многие ученые занимались дерматоглификой близнецов и спорили о степени важности показателей рельефа кожи для диагностики близнецов. Некоторые из специалистов, например Камминс (Cummins, 1930) и Райф (Rife, 1933b; Rife a. Cummins, 1943), считали их ненадежными признаками для этой цели. Их противники, наоборот, стремились доказать существенное значение этих признаков (Komai, 1928; Newman, 1930; Stocks,. 1930, 1933; Verschuer, 1933; Волоцкой, 1936; McArthur, 1938; Slater, 1953; Herrman, 1953; Wendt, 1955, и др.). В результате дактилоскопия близнецов оказалась сравнительно хорошо изученной на значительном материале в несколько сот пар близнецов и достигнуты некоторые интересные обобщения, которые говорят за то, что рельеф кожи несомненно годится для суждения о природе близнецов наравне с другими существенными для диагноза признаками. Одно из важнейших обобщений заключается в следующем «правиле», установленном Комаи и Ньюменом: в большинстве случаев отпечатки и пальцев и ладоней гомологичных рук двух однояйцевых близнецов более похожи по своим рисункам, чем правой и левой руки того же субъекта (рис. 139). Обратное, в общем, можно сказать относительно РБ (Komai, 1928; Newman, 1930, и др.).
Если подсчитать количество полосок дактилоскопических узоров и вычислить коэффициенты корреляции между правой и левой рукой какого-нибудь субъекта, взятого как из числа ОБ, так и РБ (Newman, 1930, и др.), то получится в среднем одинаковое число (r = +0.93(+/-)0.01), свидетельствующее о большой корреляции. Такова же корреляция между левыми руками ОБ одной пары и несколько меньше (r = +0.92(+/-)0.01) между правыми руками. Больше всего коэффициент корреляции (r = +0.95(+/-)0.01) между обеими руками одного из близнецов с обеими руками другого. Для РБ этот коэффициент корреляции значительно меньше (r = +0.46(+/-)0.08).
Однако было бы ошибкой думать, что внутрипарное сходство кожных узоров ОБ всегда одинаково велико и что у них всегда гомологичные пальцы имеют тождественный узор и тот же числовой показатель. Полного тождества никогда не наблюдается, и всегда можно найти известные, хотя бы небольшие, различия между узорами гомологичных пальцев, даже в случае исключительно большого сходства, как например на отпечатках рис. 141.
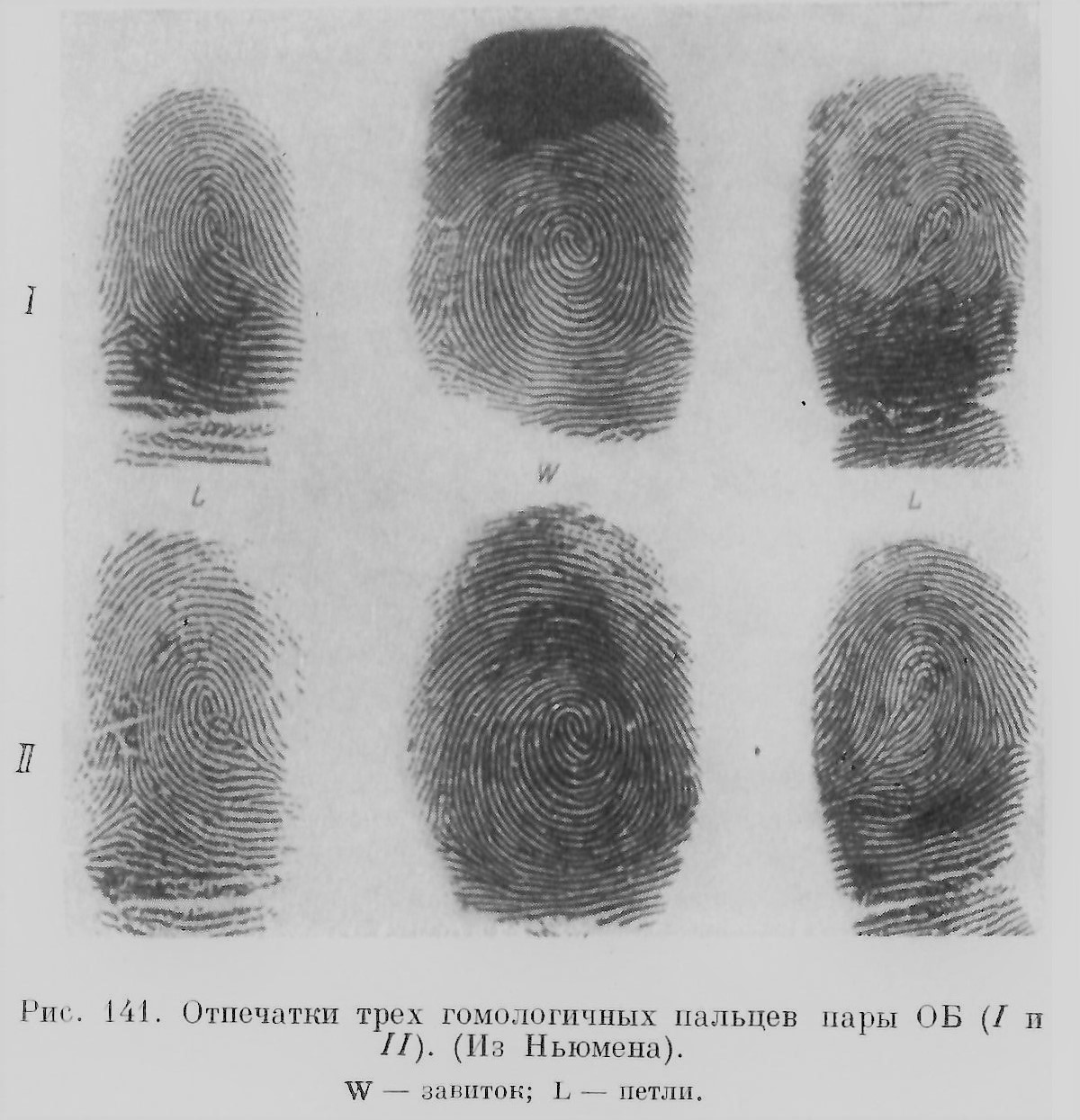 Кроме того, если коррелировать числовые показатели отдельных гомологичных пальцев у ОБ, то наблюдается известная разница между величиной коэффициента корреляции для различных пальцев, и в виде исключения коэффициент корреляции при билатеральном сравнении [14] может быть больше, чем при гомолатеральном, например для третьего пальца (Канаев, 1940а).
Фершюр (Verschuer, 1933), сравнивая разность количественных показателей дактилоскопических узоров у ОБ и двух типов РБ — одинакового пола (РБо) и разного пола (РБр), в общем подтвердил выводы Ньюмена. Эти данные Фершюра сведены в таблицу средней разницы количественных показателей трех пальцев (табл. 15).
Кроме того, если коррелировать числовые показатели отдельных гомологичных пальцев у ОБ, то наблюдается известная разница между величиной коэффициента корреляции для различных пальцев, и в виде исключения коэффициент корреляции при билатеральном сравнении [14] может быть больше, чем при гомолатеральном, например для третьего пальца (Канаев, 1940а).
Фершюр (Verschuer, 1933), сравнивая разность количественных показателей дактилоскопических узоров у ОБ и двух типов РБ — одинакового пола (РБо) и разного пола (РБр), в общем подтвердил выводы Ньюмена. Эти данные Фершюра сведены в таблицу средней разницы количественных показателей трех пальцев (табл. 15).
 Из табл. 15 видно, что различие между пальцами правой и левой руки одинаково для РБо и РБр, причем оно больше для второго пальца, чем для третьего и т. д. Такое различие имеется между правой рукой одного из ОБ и левой его партнера, и наоборот. Для РБ такая разность больше. Разность же между гомологичными руками ОБ меньше, чем между правой и левой того же субъекта, согласно «правилу» Комаи—Ньюмена.
Более простым приемом сравнения кожных узоров пальцев является подсчет числа дельт на всех десяти пальцах каждого близнеца и определение внутрипарной разности по числу дельт. На 450 парах равнополых близнецов (из которых 246 пар были ОБ, а остальные РБ), изученных Волоцким (1936), обнаруживается очень заметное большее внутрипарное сходство ОБ по сравнению с РБ, выраженное графически (рис. 142, А).
Из табл. 15 видно, что различие между пальцами правой и левой руки одинаково для РБо и РБр, причем оно больше для второго пальца, чем для третьего и т. д. Такое различие имеется между правой рукой одного из ОБ и левой его партнера, и наоборот. Для РБ такая разность больше. Разность же между гомологичными руками ОБ меньше, чем между правой и левой того же субъекта, согласно «правилу» Комаи—Ньюмена.
Более простым приемом сравнения кожных узоров пальцев является подсчет числа дельт на всех десяти пальцах каждого близнеца и определение внутрипарной разности по числу дельт. На 450 парах равнополых близнецов (из которых 246 пар были ОБ, а остальные РБ), изученных Волоцким (1936), обнаруживается очень заметное большее внутрипарное сходство ОБ по сравнению с РБ, выраженное графически (рис. 142, А).
 Аналогичные кривые получаются при подсчете числа бороздок узоров пальцев ОБ и РБ (рис. 142, Б; Newman et al., 1937).
Другой легкий способ оценки в цифрах дактилоскопических узоров был предложен сравнительно недавно (Keiter, 1950; Rife, 1952b; Wendt, 1955). Для этого отдельные типы узоров обозначаются чисто условно цифрами, например: дуга — 0, петля — 1, завиток — 2, или, учитывая переходные формы, в общем довольно редкие, берется 5 и даже 7 типов узоров, также обозначаемые цифрами — от 1 до 5 или соответственно до 7. Суммируя цифры, соответствующие узорам десяти пальцев данного субъекта, получаем одну цифру, условно, характеризующую индивидуальную картину узоров данного лица. Беря разность таких двух цифр одной пары близнецов, получаем выражение их внутрипарной разности. Применение этого приема на 100 парах ОБ и стольких же пар РБ показало,что 19% ОБ имеют разность, равную нулю, вообще же разность не более 5, тогда как среди РБ разность, равную нулю, имело лишь 5% пар, до 5 включительно — 42% РБ; остальные пары РБ этой группы имели разность до 33 (Wendt, 1955). Эти цифры также говорят о большем внутрипарном сходстве дактилоскопических рисунков у ОБ по сравнению с РБ.
Одной из последних попыток подробного изучения и практического применения к диагностике близнецов дерматоскопии является работа Мак-Артура (McArthur, 1938). Он использовал близнецовый материал Ньюмена (Newman, 1930), сравнив близнецов обоих типов между собой и с различными группами неблизнецов, одиночно рожденных людей. Полученные им данные сведены в таблицу, в которой показана средняя разность в цифрах при трояком сравнении рук (табл. 16): билатеральном, (правая и левая того же субъекта), гомолатеральиом (правая с правой и левая с левой внутрипарно) и гетеролатеральном (правая с левой и наоборот, тоже внутрипарно). Кроме близнецов, для сравнения привлекались пары родственников и пары не родственных субъектов (муж-жена, случайные пары).
Аналогичные кривые получаются при подсчете числа бороздок узоров пальцев ОБ и РБ (рис. 142, Б; Newman et al., 1937).
Другой легкий способ оценки в цифрах дактилоскопических узоров был предложен сравнительно недавно (Keiter, 1950; Rife, 1952b; Wendt, 1955). Для этого отдельные типы узоров обозначаются чисто условно цифрами, например: дуга — 0, петля — 1, завиток — 2, или, учитывая переходные формы, в общем довольно редкие, берется 5 и даже 7 типов узоров, также обозначаемые цифрами — от 1 до 5 или соответственно до 7. Суммируя цифры, соответствующие узорам десяти пальцев данного субъекта, получаем одну цифру, условно, характеризующую индивидуальную картину узоров данного лица. Беря разность таких двух цифр одной пары близнецов, получаем выражение их внутрипарной разности. Применение этого приема на 100 парах ОБ и стольких же пар РБ показало,что 19% ОБ имеют разность, равную нулю, вообще же разность не более 5, тогда как среди РБ разность, равную нулю, имело лишь 5% пар, до 5 включительно — 42% РБ; остальные пары РБ этой группы имели разность до 33 (Wendt, 1955). Эти цифры также говорят о большем внутрипарном сходстве дактилоскопических рисунков у ОБ по сравнению с РБ.
Одной из последних попыток подробного изучения и практического применения к диагностике близнецов дерматоскопии является работа Мак-Артура (McArthur, 1938). Он использовал близнецовый материал Ньюмена (Newman, 1930), сравнив близнецов обоих типов между собой и с различными группами неблизнецов, одиночно рожденных людей. Полученные им данные сведены в таблицу, в которой показана средняя разность в цифрах при трояком сравнении рук (табл. 16): билатеральном, (правая и левая того же субъекта), гомолатеральиом (правая с правой и левая с левой внутрипарно) и гетеролатеральном (правая с левой и наоборот, тоже внутрипарно). Кроме близнецов, для сравнения привлекались пары родственников и пары не родственных субъектов (муж-жена, случайные пары).
 Этицифры позволяют сделать следующие выводы. При билатеральном сравнении разница между правой и левой рукой приблизительно так же велика у обеих групп близнецов, как у случайных индивидуумов.
Этот факт несовместим с представлением, что один из ОБ сохраняет особенности правой, а другой — левой стороны первоначально единого эмбриона, т. е. противоречит вышеупомянутой теории Ньюмена.
Особенно интересно гомолатеральное сравнение рук. Внутрипарная разница у ОБ заметно меньше, чем у РБ и других сравниваемых пар. Особенно резко выделяется цифра числа полосок у ОБ по сравнению с другими парами, что заставляет Мак-Артура считать этот показатель самым тонким по сравнению с остальными тремя. Гомолатеральное сравнение ОБ с билатеральным их же и с другими типами пар говорит в пользу «правила» Комаи—Ньюмена, о котором речь была выше. Обобщенно разница между гомолатеральным сравнением ОБ, билатеральным и гомолатеральным РБ выразится в цифрах приблизительно так: 2:3:4. Но это общие средние цифры. В частных случаях внутрипарная разница у отдельных пар близнецов колеблется. Так, среди 50 пар РБ у 42 пар гомолатеральная разница была меньше, чем билатеральная, а у 8 пар — больше. Среди 50 пар РБ разница гомолатеральных рук была в 6 случаях меньше, чем право-левая разница, у 2 пар — равна и у 4 пар больше. Иногда и при сравнении двух братьев или сестер разница гомолатеральных рук оказывается меньше, чем разница между правой и левой рукой того же субъекта.
При гетеролатеральном сравнении у 7 пар ОБ обнаружилась зеркальность; у них в среднем гетеролатеральная разность была 19.5%, а гомолатеральная 25.7%, т. е. картина, обратная обычной у ОБ. У остальных ОБ гетеролатеральная разность была около 26%, т. е. близка к билатеральной разности (26.9%), и около 41—44% у РБ и братьев-сестер, в общем же несколько меньше, чем у неродных (муж-жена).
В общем у 84% РБ гомо- и гетеролатеральная разность превосходит билатеральную, тогда как у 84% ОБ либо би- и гетеролатеральная разность превышает гомолатеральную, либо гетеролатеральная разность превышает обе другие (у зеркальных пар). У 90% ОБ гомолатеральная разность была меньше 30%, а у 84% РБ одного пола была больше 30%. Отсюда Мак-Артур выводит «правило»: «Если пара близнецов имеет не больше 30% гомолатеральной разности рук, то имеется 84% вероятности, что это ОБ; если же они имеют более 30% гомолатеральной разности, то 90% вероятности, что это РБ». Таково практическое применение в диагностике близнецов полученных автором результатов. Этим приемом можно также определять членов тройни, четверни и т. д. в смысле их происхождения из одного или разных яиц. Мак-Артур считает отпечатки пальцев и ладоней достаточно точным и объективным приемом диагностики близнецов. Вероятно, это так, но следует, конечно, проверить этот метод и на другом материале и на большем числе близнецов и неблизнецов.
Сравнительно недавно отпечатки пальцев были с успехом использованы для диагноза типа близнецов психиатром Слэтером (Slater, 1953), который брал только число полосок узора и при соответствующей статистической обработке мог видеть бесспорно большее внутрипарное сходство ОБ по сравнению с РБ, особенно по общему числу полосок и коэффициенту корреляции гомологичных пальцев гомологичных рук.
Вопросом «зеркальности» рук близнецов особо занимались Райф и Камминс (Rife a. Cummins, 1943), которые также сравнивали внутрипарно не только близнецов, но и пары случайных индивидуумов. Эти авторы пришли к выводу, что зеркальность отпечатков встречается при сравнении рук случайной пары людей не менее часто, чем при внутрипарном сравнении ОБ, а потому, по их мнению, зеркальность ни в какой мере не сопряжена с процессом образования ОБ. Иначе говоря, по отпечаткам рук нельзя доказать, что один из близнецов возник из некогда правой половины исходного зародыша, а другой из левой.
В этом отношении особенно интересно сравнить отпечатки рук соединенных близнецов («сиамских»), чем и занялся Камминс (Cummins, 1934, 1936). Он использовал отпечатки пальцев 3 пар соединенных близнецов, пигопагов женского пола: Блажек, Джибб и Хилтон (последние 2 пары были тогда еще живы). Если сравнивать этих близнецов внутрипарно по типу узоров, отмечая число пар несходных пальцев при разных сравнениях, то получается следующая картина (табл. 17).
Этицифры позволяют сделать следующие выводы. При билатеральном сравнении разница между правой и левой рукой приблизительно так же велика у обеих групп близнецов, как у случайных индивидуумов.
Этот факт несовместим с представлением, что один из ОБ сохраняет особенности правой, а другой — левой стороны первоначально единого эмбриона, т. е. противоречит вышеупомянутой теории Ньюмена.
Особенно интересно гомолатеральное сравнение рук. Внутрипарная разница у ОБ заметно меньше, чем у РБ и других сравниваемых пар. Особенно резко выделяется цифра числа полосок у ОБ по сравнению с другими парами, что заставляет Мак-Артура считать этот показатель самым тонким по сравнению с остальными тремя. Гомолатеральное сравнение ОБ с билатеральным их же и с другими типами пар говорит в пользу «правила» Комаи—Ньюмена, о котором речь была выше. Обобщенно разница между гомолатеральным сравнением ОБ, билатеральным и гомолатеральным РБ выразится в цифрах приблизительно так: 2:3:4. Но это общие средние цифры. В частных случаях внутрипарная разница у отдельных пар близнецов колеблется. Так, среди 50 пар РБ у 42 пар гомолатеральная разница была меньше, чем билатеральная, а у 8 пар — больше. Среди 50 пар РБ разница гомолатеральных рук была в 6 случаях меньше, чем право-левая разница, у 2 пар — равна и у 4 пар больше. Иногда и при сравнении двух братьев или сестер разница гомолатеральных рук оказывается меньше, чем разница между правой и левой рукой того же субъекта.
При гетеролатеральном сравнении у 7 пар ОБ обнаружилась зеркальность; у них в среднем гетеролатеральная разность была 19.5%, а гомолатеральная 25.7%, т. е. картина, обратная обычной у ОБ. У остальных ОБ гетеролатеральная разность была около 26%, т. е. близка к билатеральной разности (26.9%), и около 41—44% у РБ и братьев-сестер, в общем же несколько меньше, чем у неродных (муж-жена).
В общем у 84% РБ гомо- и гетеролатеральная разность превосходит билатеральную, тогда как у 84% ОБ либо би- и гетеролатеральная разность превышает гомолатеральную, либо гетеролатеральная разность превышает обе другие (у зеркальных пар). У 90% ОБ гомолатеральная разность была меньше 30%, а у 84% РБ одного пола была больше 30%. Отсюда Мак-Артур выводит «правило»: «Если пара близнецов имеет не больше 30% гомолатеральной разности рук, то имеется 84% вероятности, что это ОБ; если же они имеют более 30% гомолатеральной разности, то 90% вероятности, что это РБ». Таково практическое применение в диагностике близнецов полученных автором результатов. Этим приемом можно также определять членов тройни, четверни и т. д. в смысле их происхождения из одного или разных яиц. Мак-Артур считает отпечатки пальцев и ладоней достаточно точным и объективным приемом диагностики близнецов. Вероятно, это так, но следует, конечно, проверить этот метод и на другом материале и на большем числе близнецов и неблизнецов.
Сравнительно недавно отпечатки пальцев были с успехом использованы для диагноза типа близнецов психиатром Слэтером (Slater, 1953), который брал только число полосок узора и при соответствующей статистической обработке мог видеть бесспорно большее внутрипарное сходство ОБ по сравнению с РБ, особенно по общему числу полосок и коэффициенту корреляции гомологичных пальцев гомологичных рук.
Вопросом «зеркальности» рук близнецов особо занимались Райф и Камминс (Rife a. Cummins, 1943), которые также сравнивали внутрипарно не только близнецов, но и пары случайных индивидуумов. Эти авторы пришли к выводу, что зеркальность отпечатков встречается при сравнении рук случайной пары людей не менее часто, чем при внутрипарном сравнении ОБ, а потому, по их мнению, зеркальность ни в какой мере не сопряжена с процессом образования ОБ. Иначе говоря, по отпечаткам рук нельзя доказать, что один из близнецов возник из некогда правой половины исходного зародыша, а другой из левой.
В этом отношении особенно интересно сравнить отпечатки рук соединенных близнецов («сиамских»), чем и занялся Камминс (Cummins, 1934, 1936). Он использовал отпечатки пальцев 3 пар соединенных близнецов, пигопагов женского пола: Блажек, Джибб и Хилтон (последние 2 пары были тогда еще живы). Если сравнивать этих близнецов внутрипарно по типу узоров, отмечая число пар несходных пальцев при разных сравнениях, то получается следующая картина (табл. 17).

 Количественные показатели узоров (число полосок) лучше демонстрируются в абсолютных числах полосок, а не в числах несходных пар пальцев (табл. 18).
Бросается в глаза, что Блажек наиболее внутрипарно различны среди этих 3 пар пигопагов, а Хилтон — наиболее похожи. При гомо-, гетеро- и билатеральном сравнении рук у Хилтон во всех 3 случаях имеется весьма небольшая разница, более или менее одинаковая, у Джибб — немного большая, но все же без существенной разницы при всех трех типах сравнения.
У Блажек тоже по разнице типов узоров нет существенного различия при этих сравнениях, но по количеству полосок близнец II явно превосходит I как по общему числу на обеих руках (94 к 78), так и для каждой руки. Поскольку статистически показано, что в населении на правой руке число полосок в общем больше, чем на левой, то можно в духе теории Ньюмена считать, что близнец II из пары Блажек — правый, a I — левый. К сожалению, когда делались отпечатки их пальцев, не было отмечено, какие отпечатки Розы, а какие Йозефы, т. е. действительно правого или левого компонента. Интересно, что число полосок на правой руке и той и другой больше, чем на левой той же особи. Из всех 3 пар, очевидно, только к Блажек применимо «правило» Комаи—Ньюмена и только в отношении числа полосок узоров, так как у них при гетеро- и билатеральном сравнении рук разность больше, чем при гомолатеральном. Таким образом, о зеркальности узоров у партнеров можно в какой-то мере говорить только в отношении Блажек. Две другие пары не обнаруживают зеркальности по отпечаткам пальцев. Казалось бы, если относительно момента их эмбрионального раздвоения можно судить по тому, что они все пигопаги, то можно было бы ждать у них и одинаковой степени зеркальности, чего, однако, по отпечаткам пальцев не видно. На этом основании, а также на основании данных Килера (Keeler, 1929) о том, что значительная зеркальность имеется у близнецов, соединенных латерально, Камминс заключает, что зеркальность зависит от иных факторов, а не от времени раздвоения, как думает Ньюмен. Камминс предполагает, что у «сиамских близнецов», как эти 3 пары пигопагов, различия партнеров, носящие характер зеркальности, зависят от различий условий утробного развития каждого из компонентов именно в связи с тем, что они соединены. Насколько это так вообще, как и какие условия влияют на те или иные свойства, в частности на узоры пальцев, — это вопрос открытый, требующий дальнейших исследований.
На дермоскопии близнецов мы остановились более подробно потому, что это признак, имеющий количественные показатели, легко и точно изучаемый, хорошо известный и несомненно ценный для диагноза типа близнецов.
Таким образом, среди различных особенностей кожи и ее производных для сравнения близнецов и выяснения их типа особенно важное значение имеют следующие: цвет кожи и волос, форма волос и узоры рельефа кожи. Далее следует расположение капилляров кожи, веснушки и другие особенности кожного покрова.
Изучение физиологии кожи дает удивительные примеры внутрипарного сходства ОБ в этой области. Известно, например, что пересадки кожи с одного субъекта на другого плохо удаются из-за несходства биохимических особенностей разных людей. Исключение представляют ОБ, у которых трансплантация кожи с одного на другого возможна, так же как при аутопластической пересадке. Такие операции делались успешно уже много раз (Bauer, 1927; Brown, 1937; Franceschetti et аl.,1949,и др.). Они свидетельствуют об исключительной близости биохимии и физиологии кожи ОБ. Глубже и подробнее это явление еще не изучено.
Внутрипарное сходство реакций кожи ОБ установлено также при различных прививках — туберкулина, противооспенной вакцины и т. п. Однако в ряде случаев обнаружена также внутрипарная дискордантность ОБ, иногда не меньшая, чем у РБ (Gedda, 1951). Чем обусловлено сходство и различие в подобных случаях, остается пока еще не выясненным.
Количественные показатели узоров (число полосок) лучше демонстрируются в абсолютных числах полосок, а не в числах несходных пар пальцев (табл. 18).
Бросается в глаза, что Блажек наиболее внутрипарно различны среди этих 3 пар пигопагов, а Хилтон — наиболее похожи. При гомо-, гетеро- и билатеральном сравнении рук у Хилтон во всех 3 случаях имеется весьма небольшая разница, более или менее одинаковая, у Джибб — немного большая, но все же без существенной разницы при всех трех типах сравнения.
У Блажек тоже по разнице типов узоров нет существенного различия при этих сравнениях, но по количеству полосок близнец II явно превосходит I как по общему числу на обеих руках (94 к 78), так и для каждой руки. Поскольку статистически показано, что в населении на правой руке число полосок в общем больше, чем на левой, то можно в духе теории Ньюмена считать, что близнец II из пары Блажек — правый, a I — левый. К сожалению, когда делались отпечатки их пальцев, не было отмечено, какие отпечатки Розы, а какие Йозефы, т. е. действительно правого или левого компонента. Интересно, что число полосок на правой руке и той и другой больше, чем на левой той же особи. Из всех 3 пар, очевидно, только к Блажек применимо «правило» Комаи—Ньюмена и только в отношении числа полосок узоров, так как у них при гетеро- и билатеральном сравнении рук разность больше, чем при гомолатеральном. Таким образом, о зеркальности узоров у партнеров можно в какой-то мере говорить только в отношении Блажек. Две другие пары не обнаруживают зеркальности по отпечаткам пальцев. Казалось бы, если относительно момента их эмбрионального раздвоения можно судить по тому, что они все пигопаги, то можно было бы ждать у них и одинаковой степени зеркальности, чего, однако, по отпечаткам пальцев не видно. На этом основании, а также на основании данных Килера (Keeler, 1929) о том, что значительная зеркальность имеется у близнецов, соединенных латерально, Камминс заключает, что зеркальность зависит от иных факторов, а не от времени раздвоения, как думает Ньюмен. Камминс предполагает, что у «сиамских близнецов», как эти 3 пары пигопагов, различия партнеров, носящие характер зеркальности, зависят от различий условий утробного развития каждого из компонентов именно в связи с тем, что они соединены. Насколько это так вообще, как и какие условия влияют на те или иные свойства, в частности на узоры пальцев, — это вопрос открытый, требующий дальнейших исследований.
На дермоскопии близнецов мы остановились более подробно потому, что это признак, имеющий количественные показатели, легко и точно изучаемый, хорошо известный и несомненно ценный для диагноза типа близнецов.
Таким образом, среди различных особенностей кожи и ее производных для сравнения близнецов и выяснения их типа особенно важное значение имеют следующие: цвет кожи и волос, форма волос и узоры рельефа кожи. Далее следует расположение капилляров кожи, веснушки и другие особенности кожного покрова.
Изучение физиологии кожи дает удивительные примеры внутрипарного сходства ОБ в этой области. Известно, например, что пересадки кожи с одного субъекта на другого плохо удаются из-за несходства биохимических особенностей разных людей. Исключение представляют ОБ, у которых трансплантация кожи с одного на другого возможна, так же как при аутопластической пересадке. Такие операции делались успешно уже много раз (Bauer, 1927; Brown, 1937; Franceschetti et аl.,1949,и др.). Они свидетельствуют об исключительной близости биохимии и физиологии кожи ОБ. Глубже и подробнее это явление еще не изучено.
Внутрипарное сходство реакций кожи ОБ установлено также при различных прививках — туберкулина, противооспенной вакцины и т. п. Однако в ряде случаев обнаружена также внутрипарная дискордантность ОБ, иногда не меньшая, чем у РБ (Gedda, 1951). Чем обусловлено сходство и различие в подобных случаях, остается пока еще не выясненным.
Скелет
О различиях формы головы близнецов, а следовательно и черепа, речь была уже выше (см. главу пятую). Вопросы развития черепа у близнецов обеих групп, окостенения различных его участков, особенностей строения и формы костей его и т. д. еще мало изучены (ср. Gedda, 1951). Развитие черепа у близнецов можно изучать путем промеров на рентгеновских снимках с последующей математической обработкой этих промеров, вычисляя, например, уклонения от определенного, условного стандарта (Patzig, 1937). При сравнении таких данных относительно черепов пары ОБ с аналогичными данными членов их семьи внутрипарное сходство черепов ОБ выступает очень отчетливо. Это направление изучения черепа мало еще разработано. Есть и другие единичные исследования отдельных деталей черепа, например пневматизации турецкого седла у близнецов, тоже говорящие о заметном сходстве ОБ (Диллон и Гуревич, 1934; Turpin, 1942, и др.). Позвоночнику близнецов посвящена капитальная монография Кюне (Kuhne, 1936). Близнецы изучались на фоне семьи, т. е. изучались и их родственники. Обследовано с помощью рентгена 53 пары ОБ, 35 пар РБ одинакового пола и 20 пар разного пола. На основании своих прежних исследований позвоночного столба у членов семей автор пришел к выводу, что существуют две основные наследственные «тенденции» в развитии позвоночника: в сторону головы и в сторону противоположную — в сторону копчика; этот вывод подтвердило и изучение близнецов. Дело в том, что «нормальное» строение позвоночника (который, как принято считать, состоит из 7 шейных, 12 грудных и т. д., всего из 33 позвонков) нередко колеблется, и происходят как бы «сдвиги» в двух названных направлениях. При таком сдвиге в сторону головы ребра нижнего грудного позвонка, 19-го от головы, оказываются в той или иной мере редуцированными, а на нижнем шейном, 7-м позвонке, наоборот, появляются ребра, правда недоразвитые (рис. 143). При обратном сдвиге — каудальном — картина противоположная: на 20-м позвонке, 1-м поясничном, появляются недоразвитые ребра, на 19-м — ребра вполне развиты и т. д. на тазе этот сдвиг к копчику может выразиться в том, что 25-й позвонок как бы выйдет из таза в поясницу, а 30-й окажется поглощенным тазом, тогда как при проявлении обратной тенденции этот позвонок будет в составе копчика и т. д. У ОБ одной пары всегда проявляется или та или другая тенденция, но она не всегда бывает одинаково сильно выражена у каждого из членов пары, как это видно, например, на рис. 143, А у ОБ с тенденцией вверх. У другой пары, взятой в качестве примера противоположной тенденции, таковая проявляется приблизительно в одинаковой степени. Среди РБ наблюдался ряд пар с противоположной тенденцией, т. е. дискордантных. Такие аномалии позвоночника, как его раздвоение (spina bifida), у близнецов изучались неоднократно (Newman а. Quisenberry, 1944; Fisher, 1952). Природа их неясна.
Скелет конечностей, кроме исследований аномалий пальцев, о которых речь будет ниже (глава двенадцатая), изучался с помощью рентгеновых лучей с точки зрения степени окостенения отдельных элементов скелета кисти рук, стопы, локтя и т. д., а также мелких различий формы этих костей в процессе их развития (Buschke, 1934, 1935; Pryor, 1939; Sontag a. Reynold, 1944, и др.). Оказывается, что и по деталям формы костей, и по порядку появления пунктов окостенения, а отчасти и по скорости процесса окостенения внутрипарное сходство у ОБ явно больше, чем у РБ. При сравнении тройни, которая состоит из двух ОБ, а третий близнец из другого яйца, указанные особенности отчетливо видны (Buschke, 1935). Этот третий близнец, по времени зачатия, в сущности, сверстник ОБ, их брат, разнояйцевый близнец обоих ОБ, заметно опережает их в процессе окостенения и имеет небольшие отличия в форме костей стопы. Сходство же ОБ чрезвычайно велико. У некоторых таких пар, и даже однояйцевых четверней, констатированы одни и те же, сравнительно редкие, особенности окостенения (Pryor, 1939).
Особенно интересны наблюдения над процессом окостенения у близнецов в динамике, когда рентгеновские снимки конечностей близнецов делались каждые 6 месяцев от 2 до 141/2 лет. Близнецы эти были однояйцевой тройней (рис. 144) с одинаковой у всех трех редкой качественной особенностью окостенения кисти обеих рук (Sontag a. Reynold, 1944). Наблюдение велось за 26 центрами окостенения. Обнаружилось, что существуют ясно наблюдаемые небольшие различия в процессе окостенения у этих близнецов.
Такие аномалии позвоночника, как его раздвоение (spina bifida), у близнецов изучались неоднократно (Newman а. Quisenberry, 1944; Fisher, 1952). Природа их неясна.
Скелет конечностей, кроме исследований аномалий пальцев, о которых речь будет ниже (глава двенадцатая), изучался с помощью рентгеновых лучей с точки зрения степени окостенения отдельных элементов скелета кисти рук, стопы, локтя и т. д., а также мелких различий формы этих костей в процессе их развития (Buschke, 1934, 1935; Pryor, 1939; Sontag a. Reynold, 1944, и др.). Оказывается, что и по деталям формы костей, и по порядку появления пунктов окостенения, а отчасти и по скорости процесса окостенения внутрипарное сходство у ОБ явно больше, чем у РБ. При сравнении тройни, которая состоит из двух ОБ, а третий близнец из другого яйца, указанные особенности отчетливо видны (Buschke, 1935). Этот третий близнец, по времени зачатия, в сущности, сверстник ОБ, их брат, разнояйцевый близнец обоих ОБ, заметно опережает их в процессе окостенения и имеет небольшие отличия в форме костей стопы. Сходство же ОБ чрезвычайно велико. У некоторых таких пар, и даже однояйцевых четверней, констатированы одни и те же, сравнительно редкие, особенности окостенения (Pryor, 1939).
Особенно интересны наблюдения над процессом окостенения у близнецов в динамике, когда рентгеновские снимки конечностей близнецов делались каждые 6 месяцев от 2 до 141/2 лет. Близнецы эти были однояйцевой тройней (рис. 144) с одинаковой у всех трех редкой качественной особенностью окостенения кисти обеих рук (Sontag a. Reynold, 1944). Наблюдение велось за 26 центрами окостенения. Обнаружилось, что существуют ясно наблюдаемые небольшие различия в процессе окостенения у этих близнецов.


 Интересно, что опережает других партнеров то один, то другой, то третий; в раннем возрасте опережал, например, самый низкорослый из трех, казалось бы отстающий в развитии (рис. 145, 146). Учитывая связь процесса окостенения с эндокринной системой и другими условиями, в частности окружающей среды, такие длительные наблюдения над динамикой развития скелета близнецов представляют значительный интерес.Но пока что этот вопрос еще мало разработан и, насколько мне известно, работа Зонтага является единственной в этом роде.
Интересно, что опережает других партнеров то один, то другой, то третий; в раннем возрасте опережал, например, самый низкорослый из трех, казалось бы отстающий в развитии (рис. 145, 146). Учитывая связь процесса окостенения с эндокринной системой и другими условиями, в частности окружающей среды, такие длительные наблюдения над динамикой развития скелета близнецов представляют значительный интерес.Но пока что этот вопрос еще мало разработан и, насколько мне известно, работа Зонтага является единственной в этом роде.
Мышечная система
Эта система изучена у близнецов сравнительно мало. В одной из последних работ (Gigas, 1941) делается попытка учесть и сравнить внутрипарное различие 10 пар ОБ, 13 пар РБ и трех троен. В целом ОБ обнаруживают 34% дискордантности, а РБ — 67%. Разумеется, это очень приблизительные цифры, говорящие лишь о большем внутрипарном сходстве мускулатуры ОБ по сравнению с РБ. Вопрос этот требует дальнейшей детальной разработки с учетом того большого значения, которое имеют для развития мускулатуры упражнения и различные факторы среды. Изучение мускулатуры двух пар зародышей ОБ показало ряд примеров как внутрипарного сходства ОБ, так и различия их, в частности асимметрии парных мышц. Чем это вызвано, объяснить не удалось (Siebert, 1937).Органы пищеварения
О губах и языке речь была уже выше. Большой интерес представляют зубы близнецов, довольно подробно и разносторонне изученные. Форма и величина коронки и корня зубов, несмотря на трудности типизации этих свойств, все же всеми исследователями этого вопроса считаются несомненно больше внутрипарно похожими у ОБ, чем у РБ. (Korkhaus, 1929, 1930, 1954: Verschuer, 1931/32; Лихтенштейн и Басина, 1936б; Gedda, 1951, и др.). Примером сравнения зубов по форме может служить следующая таблица (Bratengeier, 1934), в которой показана средняя внутрипарная разница в ширине отдельных зубов верхней челюсти, выраженная в процентах средней величины отдельного зуба (табл. 19). Мы видим, что разница в ширине резцов и клыков у ОБ заметно меньше, чем у РБ. Эти данные подтверждаются и более поздними наблюдениями (Lundstrom, 1948; Gedda, 1951, и др.).
Цвет зубов в подавляющем большинстве случаев одинаков у пары ОБ, чего нельзя сказать относительно РБ. Примером может служить табл. 20 (Korkhaus, 1930).
Мы видим, что разница в ширине резцов и клыков у ОБ заметно меньше, чем у РБ. Эти данные подтверждаются и более поздними наблюдениями (Lundstrom, 1948; Gedda, 1951, и др.).
Цвет зубов в подавляющем большинстве случаев одинаков у пары ОБ, чего нельзя сказать относительно РБ. Примером может служить табл. 20 (Korkhaus, 1930).
 Лишь в отдельных случаях наблюдается несомненное внутрипарное различие в цвете зубов ОБ (Лихтенштейн и Басина, 1936б).
Число зубов, их расположение и некоторые аномалии в этой области, а также в строении челюстей иногда обнаруживают заметное внутрипарное сходство у ОБ (рис. 147). Подобные аномалии в некоторых случаях удивительно похожи у обоих ОБ данной пары и могут служить лишним аргументом в пользу их однояйцевости (Verschuer, 1931/32; Macklin a. Moore, 1935; Лихтенштейн и Басина 1936б; Lasker a. Reynolds, 1948; Gedda, 1951; Hertel, 1955, и др.).
Лишь в отдельных случаях наблюдается несомненное внутрипарное различие в цвете зубов ОБ (Лихтенштейн и Басина, 1936б).
Число зубов, их расположение и некоторые аномалии в этой области, а также в строении челюстей иногда обнаруживают заметное внутрипарное сходство у ОБ (рис. 147). Подобные аномалии в некоторых случаях удивительно похожи у обоих ОБ данной пары и могут служить лишним аргументом в пользу их однояйцевости (Verschuer, 1931/32; Macklin a. Moore, 1935; Лихтенштейн и Басина 1936б; Lasker a. Reynolds, 1948; Gedda, 1951; Hertel, 1955, и др.).

 Многократно изучались у близнецов заболевания зубов, особенно кариес, связанный как с природными свойствами зубов, так и с возрастом, условиями среды и т. д. Ряд данных говорит о несколько большем внутрипарном сходстве ОБ по сравнению с РБ в отношении заболевания тех же зубов, развития заболевания и т. д. (рис. 148; Лихтенштейн и Басина 1936а; Nehls, 1940; Brucker, 1944, и др.).
Желудок у близнецов изучен сравнительно мало. Его форма, размеры и функции имеют большее внутрипарное сходство у ОБ, чем у РБ (Weitz, 1924, 1936; Morikawa, 1939). Но все эти особенности желудка плохо поддаются количественной оценке, что затрудняет точность сравнения. Есть указания, что кислотность желудочного сока и процесс его выделения во времени имеют известное сходство у ОБ (Glatzel, 1931). Самый процесс пищеварения, как и некоторые аномалии его, например запор, будто бы похож у ОБ (Weitz, 1936). Но все это вопросы, еще совершенно недостаточно изученные для того, чтобы можно было делать какие-нибудь обобщающие выводы. Были также попытки использовать несколько пар ОБ для выяснения некоторых вопросов пищеварения, например различия в усвояемости сырого и кипяченого молока младенцами-близнецами. Полученные здесь результаты еще незначительны.
Печень и другие органы пищеварения изучены еще меньше, чем желудок (Gedda, 1951). Морфология кишечника почти вовсе не изучена. Интересным вопросом является сходство кишечной флоры у ОБ, вероятно, сопряженное со сходством химического состава кишечного сока их (Coke, 1937), однако и этот вопрос еще очень мало изучен.
Многократно изучались у близнецов заболевания зубов, особенно кариес, связанный как с природными свойствами зубов, так и с возрастом, условиями среды и т. д. Ряд данных говорит о несколько большем внутрипарном сходстве ОБ по сравнению с РБ в отношении заболевания тех же зубов, развития заболевания и т. д. (рис. 148; Лихтенштейн и Басина 1936а; Nehls, 1940; Brucker, 1944, и др.).
Желудок у близнецов изучен сравнительно мало. Его форма, размеры и функции имеют большее внутрипарное сходство у ОБ, чем у РБ (Weitz, 1924, 1936; Morikawa, 1939). Но все эти особенности желудка плохо поддаются количественной оценке, что затрудняет точность сравнения. Есть указания, что кислотность желудочного сока и процесс его выделения во времени имеют известное сходство у ОБ (Glatzel, 1931). Самый процесс пищеварения, как и некоторые аномалии его, например запор, будто бы похож у ОБ (Weitz, 1936). Но все это вопросы, еще совершенно недостаточно изученные для того, чтобы можно было делать какие-нибудь обобщающие выводы. Были также попытки использовать несколько пар ОБ для выяснения некоторых вопросов пищеварения, например различия в усвояемости сырого и кипяченого молока младенцами-близнецами. Полученные здесь результаты еще незначительны.
Печень и другие органы пищеварения изучены еще меньше, чем желудок (Gedda, 1951). Морфология кишечника почти вовсе не изучена. Интересным вопросом является сходство кишечной флоры у ОБ, вероятно, сопряженное со сходством химического состава кишечного сока их (Coke, 1937), однако и этот вопрос еще очень мало изучен.
Органы дыхания
Гортань изучалась у близнецов в связи с их голосом. Конфигурация и размеры гортани определялись с помощью определенной методики (исследовался снаружи так называемый кадык), а также тембр и диапазон голоса с точностью до полутона. Установлено большее внутрипарное сходство ОБ по сравнению с РБ тоже и в этом отношении (Luchsinger, 1940, 1944, и др.). Аналогичные результаты получены недавно другой методикой (Gedda et al., 1955): речь близнецов записывалась, и затем близнецам предлагалось отличить свой голос от голоса своего близнеца. Опыт ставился на 104 парах близнецов обоих типов. 66% пар ОБ не могли узнать свой собственный голос; в 21 % пар ОБ только один из партнеров каждой пары узнал свой голос, и 13% пар узнали свой голос. У РБ, соответственно, проценты были: 2, 20 и 78. Этот метод Гедды осложнен призмой субъективизма испытуемых, через которую преломляется различаемый голос, призму сложную, в свою очередь требующую специального анализа. Надо заметить, что, с одной стороны, сходство голоса может быть исключительно велико также не только у близнецов, но, например, и у двух сестер или других родственников. С другой стороны, и у несомненных ОБ может быть, по-видимому, заметное различие тембра голосов, как например у известных соединенных близнецов Милли-Кристина, называвшихся «двухголовый соловей». Изучались на близнецах и некоторые аномалии техники речи, например заикание, в отношении которого также обнаружено заметное внутрипарное сходство ОБ (Luchsinger, 1940, и др.). Морфология легких близнецов, насколько мне известно, сравнительно мало изучена. Сходство формы легких установлено примерно у 2/3 пар ОБ и только у 1/4 пар РБ (Curtius u. Korkhaus, 1930). У двух пар зародышей ОБ легкие имели заметное внутрипарное различие (Siebert, 1937) и т. д. Имеется исследование емкости легких при максимальном выдохе (Werner, 1938, и др.). Получена следующая средняя разность между обоими типами близнецов при измерении выдоха с помощью аппарата Цунтца (табл. 21). Средняя внутрипарная разница у РБ более чем в 3 раза превышает таковую у ОБ. Однако в отдельных случаях и у ОБ бывает значительная внутрипарная разница, например у одной пары 320 см3, а у другой даже 440 см3. Эта разница объясняется тем, что в обоих случаях один из близнецов имел более развитую грудь благодаря соответствующим спортивным упражнениям. Разумеется, что такие внутрипарные различия носят длительный характер, поскольку в связи с разными условиями жизни пары ОБ глубоко меняется и склад их организмов.
Изучалась концентрация углекислого газа в выдыхаемом близнецами воздухе при возможно одинаковых условиях опыта: здоровом состоянии, одинаковом питании и т. д. Испытывалось по 20 пар того и другого типа (Habs, 1938). Расчет процентного уклонения от средней показал, что внутрипарная разность у группы ОБ равна 1.02, а группы РБ 3.45, т. е. у последней группы разность более чем в 3 раза превышает разность первой группы. Эти данные косвенно говорят о разной степени внутрипарной разницы обмена веществ у ОБ и РБ.
Средняя внутрипарная разница у РБ более чем в 3 раза превышает таковую у ОБ. Однако в отдельных случаях и у ОБ бывает значительная внутрипарная разница, например у одной пары 320 см3, а у другой даже 440 см3. Эта разница объясняется тем, что в обоих случаях один из близнецов имел более развитую грудь благодаря соответствующим спортивным упражнениям. Разумеется, что такие внутрипарные различия носят длительный характер, поскольку в связи с разными условиями жизни пары ОБ глубоко меняется и склад их организмов.
Изучалась концентрация углекислого газа в выдыхаемом близнецами воздухе при возможно одинаковых условиях опыта: здоровом состоянии, одинаковом питании и т. д. Испытывалось по 20 пар того и другого типа (Habs, 1938). Расчет процентного уклонения от средней показал, что внутрипарная разность у группы ОБ равна 1.02, а группы РБ 3.45, т. е. у последней группы разность более чем в 3 раза превышает разность первой группы. Эти данные косвенно говорят о разной степени внутрипарной разницы обмена веществ у ОБ и РБ.
Органы кровообращения и кровь
Формой и размерами сердца близнецов занимались сравнительно немного, пользуясь рентгеном. Относительно формы сердца, трудно поддающейся точному изучению упомянутым методом, установлено, что в среднем она внутрипарно больше похожа у ОБ, чем у РБ; однако возрастные различия при этом не учитывались, так же как не выяснялись причины различий у ОБ, кроме единичных случаев, когда, например, различие оказывалось вызванным тем, что один из партнеров занимался трудной физической работой, а другой нет (Verschuer, 1931/32). Размеры сердца близнецов также еще мало изучены. Поперечный размер сердца с учетом возраста детей от 4 до 12 лет изучался Гуревичем (1936), всего на 193 парах близнецов. Автор констатировал преобладающую роль факторов среды в изменчивости поперечного размера сердца у детей. Изучались, далее, коронарные сосуды сердца зародышей-близнецов (Tsuchiya, 1938); при этом большее внутрипарное сходство ОБ обнаруживается в небольшой мере. Другие исследования некоторых деталей анатомии сердца не дают оснований для обобщений (Gedda, 1951). Сосудистая система близнецов изучена еще меньше, чем сердце. В частности, еще очень плохо известны сосуды головного мозга, изучение которых представляет большой интерес в связи с извилинами мозга и его более тонкой структурой. Сосуды глазного дна у пары ОБ заметно различаются по своему расположению, не меньше, чем у пары РБ (Platt a. Lawton, 1956). О сосудах и капиллярах кожи говорилось выше. Что касается физиологии сердца и кровеносной системы, то она в некоторых разделах изучена несколько подробнее, чем морфология. Изучались преимущественно кровяное давление, пульс и электрокардиограммы близнецов. Кровяное давление, без точного учета возраста и условий жизни, в среднем оказалось внутрипарно почти вдвое более похожим среди ОБ по сравнению с РБ. Любопытно, что кровяное давление в детском и юношеском возрасте будто бы меняется в том смысле, что у ОБ становится внутрипарно более похожим, а у РБ, наоборот, более различным (Stocks, 1930; Verschuer, 1931/32). Примером сходства и различия кровяного давления у близнецов могут служить цифры табл. 22, где сходными считаются все пары, у которых внутрипарная разница давления не превышает 5 мм ртутного столба; все остальные случаи отнесены в рубрику дающих различие (Малкова, 1934). Процент сходства в группе ОБ в 1.7 раза больше, чем в группе РБ. Наблюдается также меньшая вариация реакции в кровяном давлении у ОБ по сравнению с РБ. Выяснение причин сходства и различия требует дальнейших исследований.
Как известно, частота пульса меняется с возрастом; на ней отражается также пол (у женщин пульс чаще). Вопрос о частоте пульса у близнецов изучен еще недостаточно, общепризнанных, обоснованных выводов нет. Наблюдается сравнительно небольшое преобладание внутрипарного сходства у ОБ по сравнению с РБ (Verschuer, 1931/32; Малкова, 1934, и др.).
Электрокардиограммы близнецов изучались сравнительно много (Weitz, 1924; Кабаков, 1930; Кабаков и Рывкин, 1924а, 1934б; Рывкин, 1936; Wise et al., 1939; Parade u. Lehmann, 1939; Kahler u. Weber, 1940; Seganti e Globbe, 1948; Gedda, 1951, и др.). По данным ряда авторов, внутрипарное сходство электрокардиограмм (ЭКГ) ОБ так велико в большинстве случаев, что по ЭКГ можно судить о типе близнецов. Примером сходства могут служить ЭКГ пары ОБ 6 лет (рис. 149); заметный контраст представляет картина внутрипарного различия у пары РБ (рис. 150). Интересно, что в ряде случаев сходство ЭКГ у взрослых ОБ было велико, несмотря на различие перенесенных болезнен и некоторых условий жизни (Кабаков и Рывкин, 1934б).
Процент сходства в группе ОБ в 1.7 раза больше, чем в группе РБ. Наблюдается также меньшая вариация реакции в кровяном давлении у ОБ по сравнению с РБ. Выяснение причин сходства и различия требует дальнейших исследований.
Как известно, частота пульса меняется с возрастом; на ней отражается также пол (у женщин пульс чаще). Вопрос о частоте пульса у близнецов изучен еще недостаточно, общепризнанных, обоснованных выводов нет. Наблюдается сравнительно небольшое преобладание внутрипарного сходства у ОБ по сравнению с РБ (Verschuer, 1931/32; Малкова, 1934, и др.).
Электрокардиограммы близнецов изучались сравнительно много (Weitz, 1924; Кабаков, 1930; Кабаков и Рывкин, 1924а, 1934б; Рывкин, 1936; Wise et al., 1939; Parade u. Lehmann, 1939; Kahler u. Weber, 1940; Seganti e Globbe, 1948; Gedda, 1951, и др.). По данным ряда авторов, внутрипарное сходство электрокардиограмм (ЭКГ) ОБ так велико в большинстве случаев, что по ЭКГ можно судить о типе близнецов. Примером сходства могут служить ЭКГ пары ОБ 6 лет (рис. 149); заметный контраст представляет картина внутрипарного различия у пары РБ (рис. 150). Интересно, что в ряде случаев сходство ЭКГ у взрослых ОБ было велико, несмотря на различие перенесенных болезнен и некоторых условий жизни (Кабаков и Рывкин, 1934б).


 Если всех близнецов по степени сходства ЭКГ разбить на три группы, то относительно большое число случаев внутрипарного сходства ОБ выступит очень отчетливо, как видно из табл. 23, охватывающей 106 пар близнецов (Parade u. Lehmann, 1939).
Несколько иные цифры, но также говорящие о большем сходстве ЭКГ у ОБ, получили Кабаков и Рывкин (1934б), изучившие 135 пар близнецов, разбив их только на две группы по сходству ЭКГ.
Наибольшее внутрипарное сходство имеет I отведение (т. е. отведение от правой и левой руки) по сравнению с другими отведениями. Не входя здесь в рассмотрение ряда интересных деталей внутрипарного сходства и различия ЭКГ близнецов, очень любопытных для суждения о физиологии и патологии сердечной мышцы, коснемся лишь зубца T,[15] который нередко бывает разной высоты у пары ОБ (рис. 151). В ряде случаев это различие зависит от размеров сердца (его поперечника и левой половины), и зубец Т выше у близнеца, имеющего большие размеры сердца. Тесная связь имеется между высотой зубца Т и некоторыми клиническими данными такими, как количество перенесенных инфекций, звучность 1-го тона, общее клиническое состояние и т. д. (Рывкин, 1936).
Если всех близнецов по степени сходства ЭКГ разбить на три группы, то относительно большое число случаев внутрипарного сходства ОБ выступит очень отчетливо, как видно из табл. 23, охватывающей 106 пар близнецов (Parade u. Lehmann, 1939).
Несколько иные цифры, но также говорящие о большем сходстве ЭКГ у ОБ, получили Кабаков и Рывкин (1934б), изучившие 135 пар близнецов, разбив их только на две группы по сходству ЭКГ.
Наибольшее внутрипарное сходство имеет I отведение (т. е. отведение от правой и левой руки) по сравнению с другими отведениями. Не входя здесь в рассмотрение ряда интересных деталей внутрипарного сходства и различия ЭКГ близнецов, очень любопытных для суждения о физиологии и патологии сердечной мышцы, коснемся лишь зубца T,[15] который нередко бывает разной высоты у пары ОБ (рис. 151). В ряде случаев это различие зависит от размеров сердца (его поперечника и левой половины), и зубец Т выше у близнеца, имеющего большие размеры сердца. Тесная связь имеется между высотой зубца Т и некоторыми клиническими данными такими, как количество перенесенных инфекций, звучность 1-го тона, общее клиническое состояние и т. д. (Рывкин, 1936).
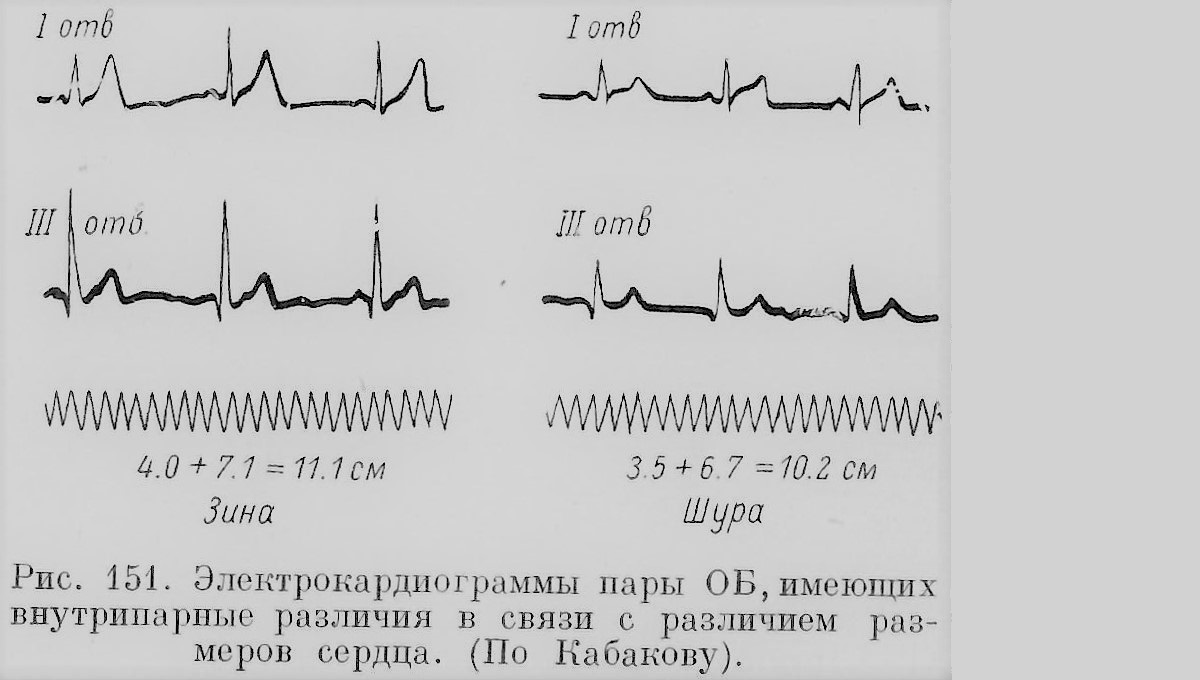 Ввиду большого внутрипарного сходства ЭКГ у ОБ изучение различий и их причин может помочь пониманию ряда зависимостей в деталях физиологии сердца.
Интересно отметить, что при situs inversus сердца зубцы при I отведении обращены вниз и зеркально в отношении к норме (рис. 152, А и Б).
Ввиду большого внутрипарного сходства ЭКГ у ОБ изучение различий и их причин может помочь пониманию ряда зависимостей в деталях физиологии сердца.
Интересно отметить, что при situs inversus сердца зубцы при I отведении обращены вниз и зеркально в отношении к норме (рис. 152, А и Б).
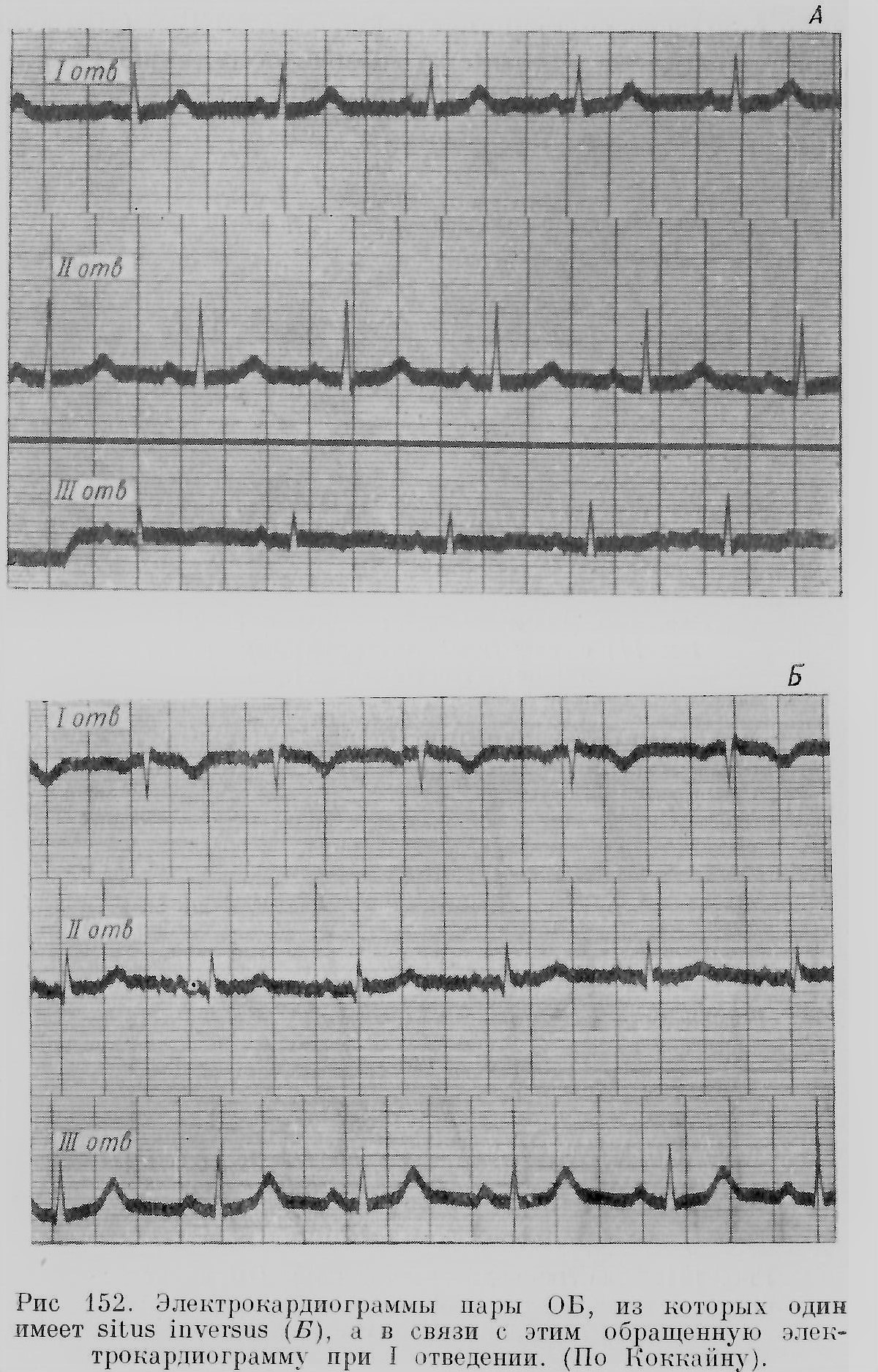 Кровь у близнецов изучена далеко не полно. Имеющиеся данные носят фрагментарный характер. Систематической картины нет (ср. Gedda, 1951). Из более новых исследований отметим исследование Тюрпена с сотрудниками (Turpin et al., 1939—1947). Установлено, что лейкоцитарная формула имеет большую впутрипарную корреляцию у ОБ (r = 0.802), чем у РБ (r = 0.419). Динамика белой крови, т. е. разных форм лейкоцитов, в связи с возрастом или одинаковыми болезнями у близнецов еще не изучена должным образом, хотя это представляет несомненный теоретический и клинический интерес. То же можно сказать относительно количества гемоглобина, биохимии крови и проч.
Особо изучалось, правда на малом числе пар, разбавление крови после питья воды — процесс очень сложный. Установлено, что внутрипарная разность у ОБ в этом отношении вдвое меньше, чем у РБ (Geyer, 1931).
Более всего внимания уделялось изучению групп и факторов крови у близнецов в связи с тем, что эти свойства крови оказались удобными для диагноза типа близнецов (Schiff u. Verschuer, 1933, и др.; ср. Gedda, 1951). Примером распределения основных групп крови у близнецов может служить следующая таблица (Verschuer, 1942).
Кровь у близнецов изучена далеко не полно. Имеющиеся данные носят фрагментарный характер. Систематической картины нет (ср. Gedda, 1951). Из более новых исследований отметим исследование Тюрпена с сотрудниками (Turpin et al., 1939—1947). Установлено, что лейкоцитарная формула имеет большую впутрипарную корреляцию у ОБ (r = 0.802), чем у РБ (r = 0.419). Динамика белой крови, т. е. разных форм лейкоцитов, в связи с возрастом или одинаковыми болезнями у близнецов еще не изучена должным образом, хотя это представляет несомненный теоретический и клинический интерес. То же можно сказать относительно количества гемоглобина, биохимии крови и проч.
Особо изучалось, правда на малом числе пар, разбавление крови после питья воды — процесс очень сложный. Установлено, что внутрипарная разность у ОБ в этом отношении вдвое меньше, чем у РБ (Geyer, 1931).
Более всего внимания уделялось изучению групп и факторов крови у близнецов в связи с тем, что эти свойства крови оказались удобными для диагноза типа близнецов (Schiff u. Verschuer, 1933, и др.; ср. Gedda, 1951). Примером распределения основных групп крови у близнецов может служить следующая таблица (Verschuer, 1942).
 Из табл. 24 видно, что дискордантных пар ОБ по группам крови нет. У РБ конкордантность тем чаще, чем чаще встречается данная группа среди населения, в данном примере — немецкого.
Полная внутрипарная конкордантность ОБ установлена также по факторам крови М и N, а также по другим факторам, в частности по резус-фактору (Rh).[16] Среди РБ встречаются, наоборот, случаи дискордантности (Strandskow a. Diederch, 1945; Schmidt, 1951, и др.). Некоторые факторы крови (Р, S и др.) еще мало изучены у близнецов. В литературе есть указание на обнаружение одной пары ОБ, дискордантной по группе крови (Panaghioton, 1938). Реальность такого случая требует проверки и должного доказательства.
Имеющиеся данные о группах и факторах крови близнецов позволяют их использовать для диагноза типа близнецов следующим образом: если пара близнецов внутрипарно дискордантна, то это явно РБ. Конкордантность ничего не доказывает, так как встречается у обоих типов близнецов.
Недавно была сделана попытка использовать группы и факторы крови, чтобы только по ним и по полу определять тип близнецов (Walsh a. Kooptzoff, 1955). Исследовалось 99 пар новорожденных близнецов. Только по полу можно из числа всех близнецов выделить 30% РБ; по полу и по группам АВО — 54% РБ; по полу, АВО и системе резус-фактора (Rh) — 65%; по полу, АВО, Rh и системе факторов MNS — 72% РБ. На основании специальных расчетов частоты встречаемости генов групп и факторов крови авторы полагают, что можно выявить почти всех РБ и тем самым диагносцировать оба основных типа — РБ и ОБ.
Интересно, что некоторые болезни крови иногда имеют большое внутрипарное сходство у ОБ. Например, пара ОБ страдала злокачественным малокровием: один заболел этим недугом в 57 лет, другой в 58; или пара других ОБ на одном и том же году, 57-м, заболела лейкемией, болезнь у обоих протекала очень похоже, и умерли они один вскоре после другого (Lotze, 1937).
Особый интерес представляет изучение гемофилии (болезни наследственной, сцепленной с полом) у ОБ, очень редко встречающейся у них. Так, например, описана пара мальчиков ОБ, из которых один страдал гемофилией, а другой был здоров (Quick a. Conway, 1949). Чем это объяснить? По-видимому, есть факторы, которые могут помешать обнаружиться гемофилии, наследуемой, как известно, связано с полом. Что это за факторы? На этот вопрос пока нет ответа.
Не изучен на близнецах еще один любопытный вопрос: предполагаемое предпочтение эктопаразитами (клопы, блохи, вши) одних хозяев другим. Насколько в этом отношении ОБ окажутся конкордантными? Это было бы тестом на биохимическое сходство крови, а также, вероятно, кожи в связи с ее запахом и проч.
Делались также попытки изучения некоторых веществ, содержащихся в крови, для сравнения метаболизма у близнецов, например количества сахара у близнецов обоих типов (Werner, 1935). Внутрипарная разность оказалась в отношении 1 : 1.3, т. е. немного больше у РБ, чем у ОБ. Результат такого исследования мало что дает, как и до сих пор сделанные попытки изучить жировой обмен, основной обмен и т. д. (Gedda, 1951). А вместе с тем все это вопросы, связанные с такой важной проблемой, как обмен веществ у близнецов.
Интересна сравнительно новая попытка Гедды (Gedda, 1951) изучить динамику обмена веществ с помощью окислительно восстановительных реакций на одном органическом веществе, трипептиде, имеющемся в крови и других тканях. Эмпирическая формула его C10H17N3SO6 (glutammil-cisteinil-glicina). Исследуя это вещество, извлеченное из крови близнецов, Гедда обнаружил удивительное внутрипарное сходство количественных показателей как окисленной, так и восстановленной фракции этого вещества, причем не только у ОБ, но и у РБ (табл. 25).
Из табл. 24 видно, что дискордантных пар ОБ по группам крови нет. У РБ конкордантность тем чаще, чем чаще встречается данная группа среди населения, в данном примере — немецкого.
Полная внутрипарная конкордантность ОБ установлена также по факторам крови М и N, а также по другим факторам, в частности по резус-фактору (Rh).[16] Среди РБ встречаются, наоборот, случаи дискордантности (Strandskow a. Diederch, 1945; Schmidt, 1951, и др.). Некоторые факторы крови (Р, S и др.) еще мало изучены у близнецов. В литературе есть указание на обнаружение одной пары ОБ, дискордантной по группе крови (Panaghioton, 1938). Реальность такого случая требует проверки и должного доказательства.
Имеющиеся данные о группах и факторах крови близнецов позволяют их использовать для диагноза типа близнецов следующим образом: если пара близнецов внутрипарно дискордантна, то это явно РБ. Конкордантность ничего не доказывает, так как встречается у обоих типов близнецов.
Недавно была сделана попытка использовать группы и факторы крови, чтобы только по ним и по полу определять тип близнецов (Walsh a. Kooptzoff, 1955). Исследовалось 99 пар новорожденных близнецов. Только по полу можно из числа всех близнецов выделить 30% РБ; по полу и по группам АВО — 54% РБ; по полу, АВО и системе резус-фактора (Rh) — 65%; по полу, АВО, Rh и системе факторов MNS — 72% РБ. На основании специальных расчетов частоты встречаемости генов групп и факторов крови авторы полагают, что можно выявить почти всех РБ и тем самым диагносцировать оба основных типа — РБ и ОБ.
Интересно, что некоторые болезни крови иногда имеют большое внутрипарное сходство у ОБ. Например, пара ОБ страдала злокачественным малокровием: один заболел этим недугом в 57 лет, другой в 58; или пара других ОБ на одном и том же году, 57-м, заболела лейкемией, болезнь у обоих протекала очень похоже, и умерли они один вскоре после другого (Lotze, 1937).
Особый интерес представляет изучение гемофилии (болезни наследственной, сцепленной с полом) у ОБ, очень редко встречающейся у них. Так, например, описана пара мальчиков ОБ, из которых один страдал гемофилией, а другой был здоров (Quick a. Conway, 1949). Чем это объяснить? По-видимому, есть факторы, которые могут помешать обнаружиться гемофилии, наследуемой, как известно, связано с полом. Что это за факторы? На этот вопрос пока нет ответа.
Не изучен на близнецах еще один любопытный вопрос: предполагаемое предпочтение эктопаразитами (клопы, блохи, вши) одних хозяев другим. Насколько в этом отношении ОБ окажутся конкордантными? Это было бы тестом на биохимическое сходство крови, а также, вероятно, кожи в связи с ее запахом и проч.
Делались также попытки изучения некоторых веществ, содержащихся в крови, для сравнения метаболизма у близнецов, например количества сахара у близнецов обоих типов (Werner, 1935). Внутрипарная разность оказалась в отношении 1 : 1.3, т. е. немного больше у РБ, чем у ОБ. Результат такого исследования мало что дает, как и до сих пор сделанные попытки изучить жировой обмен, основной обмен и т. д. (Gedda, 1951). А вместе с тем все это вопросы, связанные с такой важной проблемой, как обмен веществ у близнецов.
Интересна сравнительно новая попытка Гедды (Gedda, 1951) изучить динамику обмена веществ с помощью окислительно восстановительных реакций на одном органическом веществе, трипептиде, имеющемся в крови и других тканях. Эмпирическая формула его C10H17N3SO6 (glutammil-cisteinil-glicina). Исследуя это вещество, извлеченное из крови близнецов, Гедда обнаружил удивительное внутрипарное сходство количественных показателей как окисленной, так и восстановленной фракции этого вещества, причем не только у ОБ, но и у РБ (табл. 25).
 Далее выяснилось, что цифровые показатели этих реакций меняются изо дня в день, но всегда синхронно у одной и той же пары (табл. 26).
Но не у всех пар близнецов наблюдается такая изоритмия этих реакций. Встречаются близнецы и с несовпадающим ритмом — аллоритмические. Такая аллоритмия у ОБ бывает преходящей, вызванной вмешательством каких-либо потрясений организма (заболевания и т. п.). У РБ аллоритмия может быть и преходящей и постоянной. В материале Гедды оказалось 14.66% постоянно дискордантных в этом отношении пар РБ и ни одной нары ОБ. Гедда видит в изоритмии этих окислительно-восстановительных реакций отражение ритма обмена веществ у близнецов. Косвенное подтверждение этой мысли он находит в сходстве изменений температуры тела пары ОБ во времени (Gesell a. Thompson, 1941). Таких наблюдений, однако, еще мало.
Гедда исследовал 154 пары человеческих близнецов и поставил специальные опыты на кроликах, подтвердившие его данные, полученные на людях. Каким образом близнечество как таковое может быть причиной изоритмии у РБ — остается пока непонятным. Сам Гедда предполагает здесь явление неменделевской наследственности. Удивительна и вызывает сомнение точность совпадения цифр у близнецов: до сотых долей. Если дальнейшее исследование этого вопроса окажется в согласии с результатами Гедды, то придется признать, что он нашел интересный путь к изучению ритма метаболизма и затронул новые вопросы гемеллологии, еще ожидающие своего изучения.
Далее выяснилось, что цифровые показатели этих реакций меняются изо дня в день, но всегда синхронно у одной и той же пары (табл. 26).
Но не у всех пар близнецов наблюдается такая изоритмия этих реакций. Встречаются близнецы и с несовпадающим ритмом — аллоритмические. Такая аллоритмия у ОБ бывает преходящей, вызванной вмешательством каких-либо потрясений организма (заболевания и т. п.). У РБ аллоритмия может быть и преходящей и постоянной. В материале Гедды оказалось 14.66% постоянно дискордантных в этом отношении пар РБ и ни одной нары ОБ. Гедда видит в изоритмии этих окислительно-восстановительных реакций отражение ритма обмена веществ у близнецов. Косвенное подтверждение этой мысли он находит в сходстве изменений температуры тела пары ОБ во времени (Gesell a. Thompson, 1941). Таких наблюдений, однако, еще мало.
Гедда исследовал 154 пары человеческих близнецов и поставил специальные опыты на кроликах, подтвердившие его данные, полученные на людях. Каким образом близнечество как таковое может быть причиной изоритмии у РБ — остается пока непонятным. Сам Гедда предполагает здесь явление неменделевской наследственности. Удивительна и вызывает сомнение точность совпадения цифр у близнецов: до сотых долей. Если дальнейшее исследование этого вопроса окажется в согласии с результатами Гедды, то придется признать, что он нашел интересный путь к изучению ритма метаболизма и затронул новые вопросы гемеллологии, еще ожидающие своего изучения.
Органы выделения
Почки и другие части выделительного аппарата почти не изучались у близнецов (Gedda, 1951). Изучались, однако, различные свойства мочи, но тоже пока недостаточно. Кислотность мочи (pH) примерно также внутрипарно различна у ОБ, как и РБ (Curtius и. Korkhaus, 1930). Недавно проведено большое исследование содержания различных веществ в моче близнецов обоих типов (Berry et al., 1955). Исследование сделано на 27 парах, из которых 15 пар ОБ. У большинства моча бралась утром 3 дня подряд, но питание испытуемых было разное, что, конечно, не могло не отразиться на составе мочи. Использована была хроматографическая методика изучения мочи по Берри, и полученные данные были математически обработаны. Средняя внутрипарная разность для близнецов по четырем из изучавшихся веществ представлена в табл. 27.
Из этих цифр делается вывод, что только треонин частично находится под «генетическим контролем», поскольку внутрипарная разность у ОБ заметно меньше, чем у РБ. Чем вызвана меньшая внутрипарная разность для некоторых веществ у РБ, живущих вместе, и почему нет подобного явления у ОБ, остается неясным. Для ряда других веществ получены похожие результаты: немногие из них, как тирзин, лизин и другие, находятся под «контролем генетических факторов», относительно других веществ, как таурин, валин и т. д., этого сказать нельзя.
Железы внутренней секреции
Морфология этих желез у близнецов изучена далеко не достаточно. Тимус и щитовидная железа у двух пар зародышей ОБ имели заметное различие формы и размера (Siebert, 1937). Вместе с тем отмечается большее внутрипарное сходство развития щитовидной железы у ОБ (Curtius и. Korkhaus, 1930). Тимус, гипофиз и другие железы взрослых почти не изучались (Gedda, 1951). Патология эндокринного аппарата близнецов изучена лучше нормы (см. главу двенадцатую «Аномалии желез внутренней секреции»).Нервная система
Головной мозг близнецов еще очень мало изучен. Более ранние работы, несмотря на довольно подробное описание сходства и различия мозга близнецов, страдают тем недостатком, что их объекты неясно определены в смысле типа близнецов (Sand, 1918), — этого в начале XX в. и не могли еще сделать. Более поздних работ, когда методика определения типа близнецов была уже разработана, сравнительно мало, и касаются они лишь внешней морфологии мозга (Rossle, 1937; Siebert, 1937; Geyer, 1939; Lennox a. Gibbs, 1945, и др.). Известно, что полушария одного мозга заметно отличаются друг от друга по своему рельефу. Приблизительно такая же степень различия найдена и между мозгами пары ОБ, которые в общем внешне более похожи, чем мозги пары РБ (рис. 153, А и Б; Geyer, 1939). Подобные, в общем, отношения найдены при изучении мозга еще 71 пары близнецов (Lennox a. Gibbs, 1945). Эти данные соответствуют результатам изучения головного мозга у соединенных близнецов крупного рогатого скота (Keller u. Niedoba, 1937). Внутрипарные различия рельефа головного мозга ОБ в известной мере, вероятно, отражают различия условий их развития, которые пока еще почти не изучены. Возможно, что положение головы зародыша, кровоснабжение мозга во время развития и другие условия вызывают большую индивидуальную изменчивость мозга и его полушарий; генетически — это модификации. Гистология мозга близнецов, насколько мне известно, вовсе еще не изучалась.
 Если головной мозг близнецов еще мало изучен, то особенности спинного мозга, симпатической и периферической нервной системы почти вовсе не исследованы (Gedda, 1951). Имеются краткие сведения об относительно значительном сходстве больших нервных стволов у двух пар зародышей ОБ и заметных различиях нервных сплетений (plexus brachialis и др., — Siebert, 1937). Лишь в одной работе дается довольно подробное описание некоторых кожных нервов рук 11 пар мертворожденных близнецов (Kadanoff, 1939). Тип близнецов определялся по оболочкам. Различие между нервами гомологичных рук пары ОБ по крайней мере так же велико, как между руками одного и того же индивидуума (рис. 137). Эта изменчивость, очевидно, говорит об очень значительной роли условий развития нервов; но это еще вовсе не изученная область. Каданов изучал и особенности ветвления вен верхних конечностей близнецов. Связи между особенностями вен и нервов он не устанавливает, но констатирует большую изменчивость вен по сравнению с нервами. О его данных относительно вен речь идет в другом месте («Кожа»).
Физиология нервной системы близнецов изучена еще очень мало. Имеются лишь отдельные исследования или небольшие серии исследований некоторых вопросов этой большой области.
Неоднократно предпринимались исследования биотоков мозга близнецов (Davis Н. a. Davis, 1936; Lennox a. Gibbs, 1945, и др.). Так, из 55 пар ОБ 85% имели «идентичные» электроэнцефалограммы (рис. 154), 4% — дискордантные и 11% — «сомнительные» в смысле сходства. Из 19 пар РБ 95% имели внутрипарно неодинаковые электроэнцефалограммы и 5% похожие. «Типовое» внутрипарное сходство ОБ напоминает здесь аналогичную картину с электрокардиограммами. Леннокс считает, что электроэнцефалограммы годятся для определения одноянцевости близнецов. Биотоки мозга близнецов еще мало изучены, они совсем не исследовались в связи с изучением высшей нервной деятельности, а также электрофизиологии двигательных реакций. При изучении эпилепсии электроэнцефалограммы казались очень полезными. Нет еще систематических данных о биотоках мозга близнецов в связи с его асимметрией, что было бы интересно исследовать. В ряде функций нервной системы асимметрия мозга обнаруживается очень ясно, особенно в разнице работы рук, фило- и онтогенетически, вероятно, связанной с речевой функцией.
Если головной мозг близнецов еще мало изучен, то особенности спинного мозга, симпатической и периферической нервной системы почти вовсе не исследованы (Gedda, 1951). Имеются краткие сведения об относительно значительном сходстве больших нервных стволов у двух пар зародышей ОБ и заметных различиях нервных сплетений (plexus brachialis и др., — Siebert, 1937). Лишь в одной работе дается довольно подробное описание некоторых кожных нервов рук 11 пар мертворожденных близнецов (Kadanoff, 1939). Тип близнецов определялся по оболочкам. Различие между нервами гомологичных рук пары ОБ по крайней мере так же велико, как между руками одного и того же индивидуума (рис. 137). Эта изменчивость, очевидно, говорит об очень значительной роли условий развития нервов; но это еще вовсе не изученная область. Каданов изучал и особенности ветвления вен верхних конечностей близнецов. Связи между особенностями вен и нервов он не устанавливает, но констатирует большую изменчивость вен по сравнению с нервами. О его данных относительно вен речь идет в другом месте («Кожа»).
Физиология нервной системы близнецов изучена еще очень мало. Имеются лишь отдельные исследования или небольшие серии исследований некоторых вопросов этой большой области.
Неоднократно предпринимались исследования биотоков мозга близнецов (Davis Н. a. Davis, 1936; Lennox a. Gibbs, 1945, и др.). Так, из 55 пар ОБ 85% имели «идентичные» электроэнцефалограммы (рис. 154), 4% — дискордантные и 11% — «сомнительные» в смысле сходства. Из 19 пар РБ 95% имели внутрипарно неодинаковые электроэнцефалограммы и 5% похожие. «Типовое» внутрипарное сходство ОБ напоминает здесь аналогичную картину с электрокардиограммами. Леннокс считает, что электроэнцефалограммы годятся для определения одноянцевости близнецов. Биотоки мозга близнецов еще мало изучены, они совсем не исследовались в связи с изучением высшей нервной деятельности, а также электрофизиологии двигательных реакций. При изучении эпилепсии электроэнцефалограммы казались очень полезными. Нет еще систематических данных о биотоках мозга близнецов в связи с его асимметрией, что было бы интересно исследовать. В ряде функций нервной системы асимметрия мозга обнаруживается очень ясно, особенно в разнице работы рук, фило- и онтогенетически, вероятно, связанной с речевой функцией.
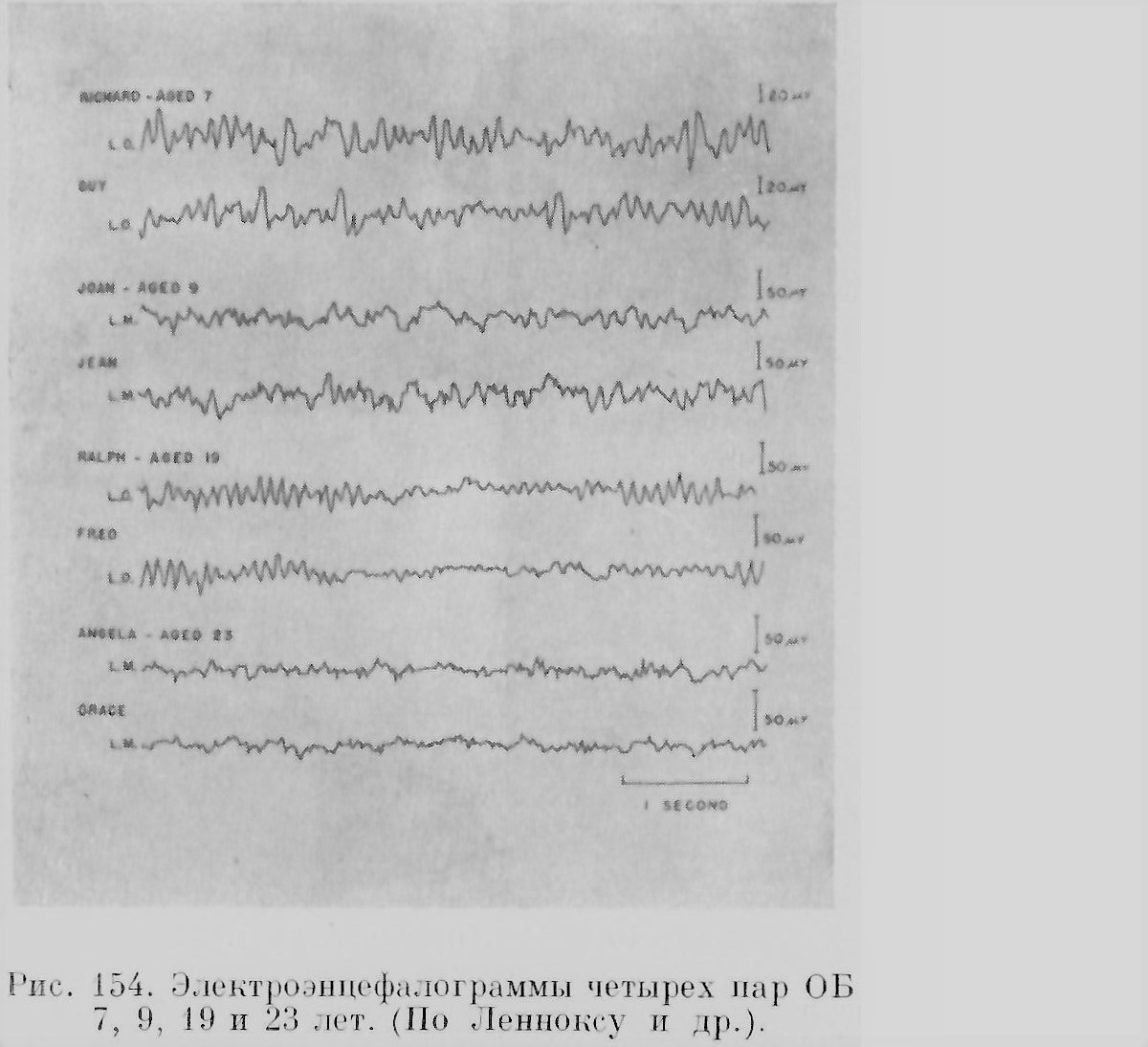
Физиология анализаторов
Из так называемых пяти «чувств» человека у близнецов кожная рецепция (осязание и проч.) и обоняние, в сущности, вовсе не изучались. Сравнительно немного изучался вкус, очень мало слух и больше всего зрение. Одним из тестов на вкус до недавнего времени служил порошок фенил-тио-карбамида (РТС), который одними лицами оценивался как горькое вещество, а другими как безвкусное. Считалось, что та или иная оценка РТС — наследственное свойство. У значительного числа ОБ оценка этого вещества совпадала. Однако встречались пары ОБ, дискордантные в этом отношении (Ардашников и др., 1936а, 1936б). Недавно установлены новые случаи дискордантных пар по определению вкуса РТС (lее, 1956). При разной степени разведения раствора РТС встречались пары ОБ, дискордантные в оценке вкуса его в убывающем числе в соответствии с возрастающей слабостью раствора. Внутрипарная конкордантность РБ была заметно меньше. И с помощью других веществ (сода, хинин и т. д.) испытывался вкус близнецов, при этом также установлено большее внутрипарное сходство ОБ по сравнению с РБ (Habs, 1938). Специально изучался вкус пятерни Дионн (Ford a. Mason, 1941). Все пять признаны конкордантными. Слуховой анализатор до сих пор остается почти не изученным у близнецов. Сравнительно давняя работа Макферлана (Macfarlan, 1927) констатирует с помощью аудиометра очень большое сходство у одной пары взрослых ОБ в восприятии звуков: кривые обоих носят аналогичный характер. В свете павловского учения об анализаторах изучение близнецов в этом отношении может дать много интересного. Физиология зрения изучалась с разных сторон, но преимущественно интересовались рефракцией (Waardenburg, 1930; Huber, 1931; Holste, 1940, и др.). Внутрипарная конкордантность ОБ оказалась значительно больше, чем РБ. Так, Варденбург получил следующую картину дискордантности среди ОБ.Разница в диоптриях 0.25__0.5__0.75__1__1.25__1.5__1.75__1.9 Число случаев 44___26___15___8___11___1____0_____9Большинство пар ОБ, как видно, имеет очень небольшую впутрипарную разницу рефракции. У РБ одинакового пола внутрипарная разница достигает 4.2—4.9 диоптрии, бывает даже больше (Glatzel, 1931). Однако существуют отдельные случаи очень резкого внутрипарного различия рефракции и у ОБ, причины которых пока не выяснены (ср. Orth, 1954, и др.). Адаптация к темноте, как известно, проявляется с заметными индивидуальными различиями у разных субъектов. Исследование адаптации у 20 пар ОБ и стольких же РБ одинакового пола показало, что внутрипарная разность у ОБ заметно меньше, чем у РБ (рис. 155). Разница может еще зависеть от обмена веществ, в частности от содержания в пище витаминов А и D. Экспериментальное изучение пар ОБ, в которых один из близнецов являлся бы контрольным, могло бы помочь в изучении дозировок витаминов для адаптации к темноте и влияния других условий.
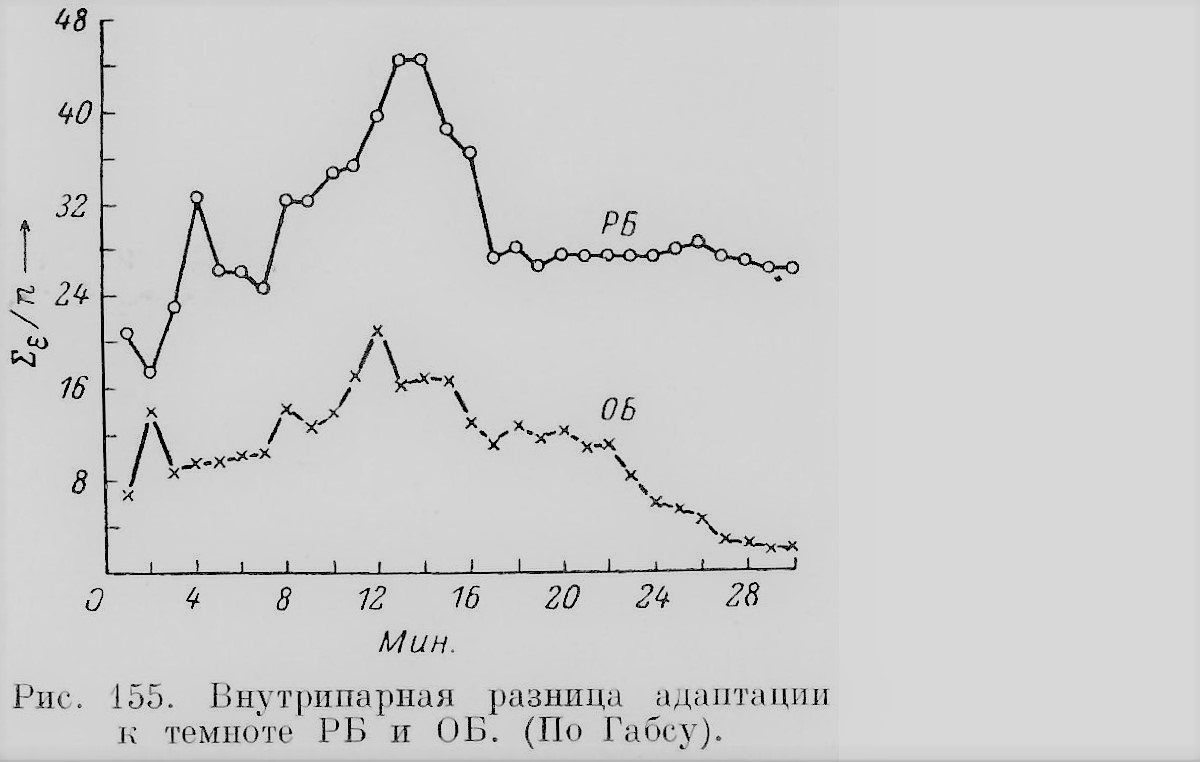 С психологических позиций экспериментально изучались явления зрительных следов, а также так называемые эйдетические образы (воспроизведение более давних зрительных следов). Внутрипарное сходство ОБ и в этой области оказалось в общем больше, чем РБ (Smith, 1949). Вероятно, здесь речь идет о функциональном сходстве мозгового конца зрительного анализатора.
Зрачковый рефлекс также оказался внутрипарно более похожим у ОБ (Lowenstein, 1939).
Интересно, что даже характер движения глаз при чтении у ОБ обнаружил явное внутрипарное сходство (Jones а. Morgan, 1942), как это видно из рис. 156, где представлены фотографии луча света (выполненные с помощью офталмографа), отраженного с роговицы глаз читающего субъекта. При математической обработке таких записей можно вычислить коэффициенты корреляции для пар ОБ, РБ и пар из случайных людей.Получаются следующие средние цифры для этих коэффициентов, соответственно, 0.53, 0.43 и 0.1. Другой методикой тоже получены до известной степени аналогичные данные, показывающие, что большее внутрипарное сходство нельзя объяснить одной «привычкой» (Morgan, 1939).
С психологических позиций экспериментально изучались явления зрительных следов, а также так называемые эйдетические образы (воспроизведение более давних зрительных следов). Внутрипарное сходство ОБ и в этой области оказалось в общем больше, чем РБ (Smith, 1949). Вероятно, здесь речь идет о функциональном сходстве мозгового конца зрительного анализатора.
Зрачковый рефлекс также оказался внутрипарно более похожим у ОБ (Lowenstein, 1939).
Интересно, что даже характер движения глаз при чтении у ОБ обнаружил явное внутрипарное сходство (Jones а. Morgan, 1942), как это видно из рис. 156, где представлены фотографии луча света (выполненные с помощью офталмографа), отраженного с роговицы глаз читающего субъекта. При математической обработке таких записей можно вычислить коэффициенты корреляции для пар ОБ, РБ и пар из случайных людей.Получаются следующие средние цифры для этих коэффициентов, соответственно, 0.53, 0.43 и 0.1. Другой методикой тоже получены до известной степени аналогичные данные, показывающие, что большее внутрипарное сходство нельзя объяснить одной «привычкой» (Morgan, 1939).
 В общем, как видно из изложенного, физиология анализаторов у близнецов изучена еще мало, а в связи с высшей нервной деятельностью и вовсе не изучалась.
В общем, как видно из изложенного, физиология анализаторов у близнецов изучена еще мало, а в связи с высшей нервной деятельностью и вовсе не изучалась.
Физиология высшей нервной деятельности
Физиология высшей нервной деятельности близнецов еще очень мало исследована (Канаев, 1934—1959; Анохин, 1939; Молчанова-Сеничева, 1941; Алексеева, 1941; Алексеева и Островская, 1953). Как известно, она изучается методом условных рефлексов. До сих пор работа велась лишь на младенцах первых месяцев жизни и на детях школьного возраста. В связи с возрастом детей методика применялась различная. Условные рефлексы у ОБ первых месяцев жизни изучались следующим методом: условным раздражителем служило покачивание ребенка вверх-вниз с помощью специального приспособления, а безусловным раздражителем (подкреплением ) ритмическое дуновение в глаза струей воздуха одинаковой силы из резинового баллона, находящегося всегда на одном и том же расстоянии от лица ребенка, что и вызывало рефлекторное закрывание глаз. Покачивание начиналось за 2—3 секунды до подкрепления. Совместное действие раздражителей длилось 15 секунд. Во время опыта применялось 6—10 сочетаний. Опыты обычно начинались на первом месяце жизни при одинаковых условиях. У двух из первых трех пар условный рефлекс появился одновременно, у третьей пары — с разницей в 3 дня. Число сочетаний, необходимых для получения условного рефлекса, было неодинаковым для разных пар и притом довольно значительным, например 59 сочетании и т. д. У близнецов по сравнению с нормальными детьми-одиночками условный рефлекс образуется с опозданием около месяца. Незначительные внутрипарные различия констатированы при выработке дифференцировки путем изменения направления покачивания ребенка. Но разные пары вырабатывали дифференцировку очень различно. Так, одна пара дифференцировала новое раздражение сразу, а другая на 23—25-м опыте, с известной внутрипарной разницей. При выработке условного тормоза, а затем и запаздывающего условного рефлекса также наблюдалось явное внутрипарное сходство близнецов с значительным различием между парами. Замечательный внутрипарный параллелизм в развитии близнецов обнаружен при регистрации у них первого появления ряда различных реакций: первой улыбки, первого толкания погремушки, поварачивания головы в сторону звука, хватания предмета и т. д. (Молчанова-Сеничева, 1941). К сожалению, это интересное исследование было прервано войной, а по окончании ее не возобновлено. Изучение физиологии высшей нервной деятельности и надо начинать, как в упомянутом исследовании, именно с первого месяца жизни, ведя параллельно с экспериментальной работой возможно разностороннее систематическое наблюдение за развитием близнецов, их заболеваниями и т. д. По мере роста детей должны меняться и методы эксперимента: применяться секреторно-двигательная методика, разные варианты двигательной и особенно тщательно должно изучаться развитие речи всеми доступными способами. Только многолетнее исследование развития высшей нервной деятельности с первого месяца жизни даст возможность достаточно полно изучить тип нервной системы близнецов и выяснить те условия, от которых зависит образование различных индивидуальных особенностей «склада» высшей нервной деятельности, «характера» каждого субъекта, и, следовательно, причины тех внутрипарных различий высшей нервной деятельности, которые наблюдаются в некоторых случаях в очень контрастной форме. При изучении высшей нервной деятельности школьников применялась преимущественно секреторно-двигательная методика Красногорского. Регистрировалось количество слюны в каплях под влиянием действия безусловных и условных раздражителей и двигательная реакция рта. Для изучения некоторых специальных рефлексов, как условный рефлекс на время, применялась двигательная методика на электроключах. Безусловный слюнный рефлекс на определенную порцию пищи (клюква в сахаре) за определенный отрезок времени после получения пищи, измеренный за ряд опытных дней путем подсчета капель слюны, был внутрипарно более похож у ОБ, чем у РБ, как это видно из табл. 28, где цифры обозначают среднюю внутрипарную разность в каплях слюны (Канаев, ,1939б). В группе ОБ все дети, кроме пары См., девочки, в группе РБ первые две пары девочки, третья пара (Богд.) разного пола. Интересно отметить, что величина внутрипарной разницы изо дня в день колеблется и зависит не только от множества случайных меняющихся обстоятельств, как например степень голода или насыщения, но также от индивидуального отношения ребенка к данному роду пищи, что является уже относительно стойко сложившейся реакцией данного субъекта. Например, один из близнецов См. (рис. 1) «не любил» клюкву, и потому безусловная секреция слюны у него на клюкву была меньше, чем у его близнеца, охотно евшего клюкву в сахаре. Чем вызвана была «нелюбовь» к клюкве у этого близнеца См., выяснить не удалось; вероятно, какие-то обстоятельства жизни послужили причиной возникновения данного различия в таком «прирожденном» рефлексе, каким является безусловный слюнный рефлекс (Канаев, 1940б).
Условный рефлекс у большинства ОБ возникал внутрипарно одновременно или почти одновременно, но у разных пар в разные дни, например у пары Лоб. на 5-й опытный день, у пары См. — на 3-й, у пары Кудр. — на 7-й и т. д. У пар РБ внутрипарная разница была от 2 до 5 дней. Величина условного рефлекса, измеренного числом капель слюны, была в ряде случаев очень различна у ОБ. Такое различие иногда проявлялось месяцы и годы. Этим обнаруживалось заметное различие «склада» высшей нервной деятельности, «характеров» двух близнецов одной пары. Например, у ОБ Наташи и Оксаны Гапч. за длительный срок (50 опытных дней) в опытах по дифференцировке двух разных темпов метронома, из которых один подкреплялся шоколадом, а другой не подкреплялся, оказалось, что средний уровень возбудимости у Оксаны выше, чем у Наташи, тогда как торможение у них почти одинаковое и у обеих не полное (рис. 157). В общем же дифференцировка у Оксаны примерно на 10% лучше, чем у Наташи (Канаев, 1954). Аналогичную разницу, еще более резко выраженную в силу некоторого патологического характера этой пары, обнаружила пара ОБ — Тамара и Нина Ин. (Канаев, 1937). У этой пары выявилась также большая разница при испытании с «законом силы» (усиление реакции с усилением раздражителя). У одной из девочек, Нины, на сильный звонок выделялось заметно большее число капель, чем на слабый, тогда как у другой не было разницы, как видно из средних чисел количества слюны за 30 опытных дней (табл. 29).
В группе ОБ все дети, кроме пары См., девочки, в группе РБ первые две пары девочки, третья пара (Богд.) разного пола. Интересно отметить, что величина внутрипарной разницы изо дня в день колеблется и зависит не только от множества случайных меняющихся обстоятельств, как например степень голода или насыщения, но также от индивидуального отношения ребенка к данному роду пищи, что является уже относительно стойко сложившейся реакцией данного субъекта. Например, один из близнецов См. (рис. 1) «не любил» клюкву, и потому безусловная секреция слюны у него на клюкву была меньше, чем у его близнеца, охотно евшего клюкву в сахаре. Чем вызвана была «нелюбовь» к клюкве у этого близнеца См., выяснить не удалось; вероятно, какие-то обстоятельства жизни послужили причиной возникновения данного различия в таком «прирожденном» рефлексе, каким является безусловный слюнный рефлекс (Канаев, 1940б).
Условный рефлекс у большинства ОБ возникал внутрипарно одновременно или почти одновременно, но у разных пар в разные дни, например у пары Лоб. на 5-й опытный день, у пары См. — на 3-й, у пары Кудр. — на 7-й и т. д. У пар РБ внутрипарная разница была от 2 до 5 дней. Величина условного рефлекса, измеренного числом капель слюны, была в ряде случаев очень различна у ОБ. Такое различие иногда проявлялось месяцы и годы. Этим обнаруживалось заметное различие «склада» высшей нервной деятельности, «характеров» двух близнецов одной пары. Например, у ОБ Наташи и Оксаны Гапч. за длительный срок (50 опытных дней) в опытах по дифференцировке двух разных темпов метронома, из которых один подкреплялся шоколадом, а другой не подкреплялся, оказалось, что средний уровень возбудимости у Оксаны выше, чем у Наташи, тогда как торможение у них почти одинаковое и у обеих не полное (рис. 157). В общем же дифференцировка у Оксаны примерно на 10% лучше, чем у Наташи (Канаев, 1954). Аналогичную разницу, еще более резко выраженную в силу некоторого патологического характера этой пары, обнаружила пара ОБ — Тамара и Нина Ин. (Канаев, 1937). У этой пары выявилась также большая разница при испытании с «законом силы» (усиление реакции с усилением раздражителя). У одной из девочек, Нины, на сильный звонок выделялось заметно большее число капель, чем на слабый, тогда как у другой не было разницы, как видно из средних чисел количества слюны за 30 опытных дней (табл. 29).

 Такие длительно наблюдаемые индивидуальные особенности реакций обнаруживаются как бы на фоне общих обоим близнецам типовых свойствах нервной системы, например: у близнецов Гапч. на фоне известной неуравновешенности — преобладание возбуждения над торможением, у близнецов Ин. — на фоне слабого типа обоих и т. д.
Подвижность нервных процессов, пока еще мало изученная у близнецов, оказалась очень похожей у одной пары ОБ (Канаев, 1941). Условный рефлекс на время, изучавшийся путем подачи ребенком сигнала электроключом через интервалы в 3, 6 и 12 минут, после которых давалось подкрепление (конфета), проявлялся у ОБ той же пары неодинаково в смысле точности дачи сигнала. Однако процент досрочно данных сигналов и даже степень неточности этих сигналов при интервале в 3 минуты у одной пары ОБ, мальчиков 14 лет Сав., оказались удивительно похожими. Физиологический механизм отсчета времени человеком еще мало изучен, особенно же у близнецов, и потому эти явления пока еще трудно объяснить.
Попытка исследовать, при каком темпе по метроному эта же пара ОБ выполняет определенный двигательный стереотип обеими руками на электроключах с наименьшим числом ошибок, показала значительное внутрипарное сходство этих ОБ: для обоих, в отличие от некоторых других детей, оптимальным оказался один и тот же темп — максимально скорый.
Большой интерес представляют внутрипарные различия физиологии речи близнецов, которые тесно связаны с особенностями типа и «склада» нервной системы. Только недавно началось исследование речи ОБ новой методикой, позволяющей точно регистрировать темп речевого потока и силу звуков произносимых слов.
Если предполагать, что у пары ОБ должны быть какие-нибудь общие типовые свойства нервной системы в силу общности их наследственности, то различия «склада» нервной системы, «характеров», надо, по-видимому, искать прежде всего в особенностях развития каждого из близнецов и в тех факторах среды, которые влияют на развитие. «Пластичность» нервной системы в этой связи, как указывал И. П. Павлов, очень велика. Развитие индивидуальных особенностей нервной системы близнецов в зависимости от факторов среды, начиная с утробной жизни, пока еще систематически не изучалось. Это дело будущего. В настоящее время мы можем только в порядке более или менее вероятной догадки предполагать существование известной зависимости определенных свойств нервной системы от определенных условий развития того или иного близнеца. Так, например, между Наташей и Оксаной Гапч., о которых речь была выше, существует несомненное, ясно наблюдаемое различие поведения в силу различия характеров их. Наташа, более деятельная и инициативная в играх, домашних работах и т. д., занимает ведущую роль, а Оксана, более робкая и пассивная, подчиняется Наташе, следует за ней. Это основное различие имеет свою длинную и сложную историю. Оксана при рождении весила почти на килограмм меньше, чем Наташа, и притом была синюшная и очень слабая, тогда как Наташа находилась в относительно благополучном состоянии. Это различие, обусловленное менее благоприятными обстоятельствами утробной жизни Оксаны, усилилось еще оттого, что в раннем детстве Оксана очень тяжело болела корью, а Наташа перенесла ее сравнительно легко. Оксана длительное время немного отставала в развитии и к 16 годам так и осталась несколько ниже Наташи ростом. Так как Наташа и родилась на 50 минут раньше Оксаны, то в связи с тем, что она опережала Оксану в развитии, ее стали дома считать «старшей», а Оксану — «младшей». Это внутрисемейное отношение между близнецами постепенно развивалось и крепло. В результате оно глубоко запечатлелось на всем «складе» высшей нервной деятельности близнецов, выразилось в том различии «характеров», которое выше отмечалось (Канаев, 1954).
Нечто подобное пришлось наблюдать на паре ОБ Ин. Одна из девочек этой пары — Нина — в раннем детстве имела воспаление уха, в связи с чем у нее возникла кривошея, сопровождавшаяся и некоторым отставанием в развитии по сравнению с ее близнячкой Тамарой. Последняя в этой паре оказалась в роли «ведущей», а Нина — «подчиненной», что вызвало целый ряд различий «склада» их высшей нервной деятельности (Канаев, 1937).
Как видно из этих первых попыток изучения физиологии высшей нервной деятельности, исследование ее в течение возможно длительного периода развития близнецов может дать ряд ценных сведении для понимания природы типа высшей нервной деятельности и ее «склада» в связи с условиями жизни, проливая новый свет на те материалы, которые получила психология по онтогенезу психики у близнецов (см. главу одиннадцатую).
Результатов исследования физиологии высшей нервной деятельности у соединенных близнецов (двух пар), изучавшихся в Москве, опубликовано еще очень мало (Анохин, 1939; Алексеева, 1941; Алексеева и Островская, 1953).
Физиология сна близнецов исследована еще мало. Павловская теория сна получила интересное подтверждение благодаря изучению двух пар соединенных близнецов Иры-Гали, родившихся в Москве в 1938 г. (Анохин, 1939; Алексеева, 1941). Многократно наблюдалось, что одна из близнячек спала, в то время как вторая находилась в состоянии бодрствования: смеялась, плакала и т. п. (рис. 53).
Кровеносная система этих близнецов сообщалась; следовательно, если бы сон зависел от содержания определенных веществ в крови (гуморальная теория сна), то близнецы засыпали бы всегда одновременно. Объяснение сна тормозным состоянием коры и нижележащих отделов мозга, как это делал И. П. Павлов, вполне соответствует наблюдавшейся на этой паре близнецов картине сна. Аналогичные данные получены и на второй паре соединенных близнецов (Алексеева и Островская, 1953). Интересно, что внутрипарные различия дыхания и частоты пульса этих близнецов, наблюдаемые во время их бодрствования, при одновременном глубоком сне в значительной мере исчезают.
Наблюдениями над сном целой серии пар ОБ и РБ установлено, что многие особенности сна — поза во время сна, глубина сна, его расстройства и т. д. — чаще конкордантны у ОБ, чем у РБ (Geyer, 1937). Однако такого рода исследование требует более точной постановки изучения физиологии сна, чем это делалось до сих нор, и привлечения эксперимента.
Как известно, во время сна моторная хронаксия у человека несколько увеличивается, т. е. возбудимость его тканей снижается в связи с сонным торможением, что обнаруживается в замедлении их реакции. Была сделана попытка, насколько мне известно, пока что единственная, исследовать моторную хронаксию у пары ОБ на первом году их жизни (Короткин и Крышова, 1948). Изучалась хронаксия флексоров и экстензоров пальцев руки. Обнаружено известное сходство динамики хронаксии у обоих близнецов, отличающее их от сверстников. Средние величины (из 100 отдельных измерений) хронаксии флексоров и экстензоров в бодрствующем и сонном состоянии оказались у близнецов очень схожими с таковыми у пары сверстников (рис. 158). К сожалению, это интересное исследование не было продолжено и развито.
Такие длительно наблюдаемые индивидуальные особенности реакций обнаруживаются как бы на фоне общих обоим близнецам типовых свойствах нервной системы, например: у близнецов Гапч. на фоне известной неуравновешенности — преобладание возбуждения над торможением, у близнецов Ин. — на фоне слабого типа обоих и т. д.
Подвижность нервных процессов, пока еще мало изученная у близнецов, оказалась очень похожей у одной пары ОБ (Канаев, 1941). Условный рефлекс на время, изучавшийся путем подачи ребенком сигнала электроключом через интервалы в 3, 6 и 12 минут, после которых давалось подкрепление (конфета), проявлялся у ОБ той же пары неодинаково в смысле точности дачи сигнала. Однако процент досрочно данных сигналов и даже степень неточности этих сигналов при интервале в 3 минуты у одной пары ОБ, мальчиков 14 лет Сав., оказались удивительно похожими. Физиологический механизм отсчета времени человеком еще мало изучен, особенно же у близнецов, и потому эти явления пока еще трудно объяснить.
Попытка исследовать, при каком темпе по метроному эта же пара ОБ выполняет определенный двигательный стереотип обеими руками на электроключах с наименьшим числом ошибок, показала значительное внутрипарное сходство этих ОБ: для обоих, в отличие от некоторых других детей, оптимальным оказался один и тот же темп — максимально скорый.
Большой интерес представляют внутрипарные различия физиологии речи близнецов, которые тесно связаны с особенностями типа и «склада» нервной системы. Только недавно началось исследование речи ОБ новой методикой, позволяющей точно регистрировать темп речевого потока и силу звуков произносимых слов.
Если предполагать, что у пары ОБ должны быть какие-нибудь общие типовые свойства нервной системы в силу общности их наследственности, то различия «склада» нервной системы, «характеров», надо, по-видимому, искать прежде всего в особенностях развития каждого из близнецов и в тех факторах среды, которые влияют на развитие. «Пластичность» нервной системы в этой связи, как указывал И. П. Павлов, очень велика. Развитие индивидуальных особенностей нервной системы близнецов в зависимости от факторов среды, начиная с утробной жизни, пока еще систематически не изучалось. Это дело будущего. В настоящее время мы можем только в порядке более или менее вероятной догадки предполагать существование известной зависимости определенных свойств нервной системы от определенных условий развития того или иного близнеца. Так, например, между Наташей и Оксаной Гапч., о которых речь была выше, существует несомненное, ясно наблюдаемое различие поведения в силу различия характеров их. Наташа, более деятельная и инициативная в играх, домашних работах и т. д., занимает ведущую роль, а Оксана, более робкая и пассивная, подчиняется Наташе, следует за ней. Это основное различие имеет свою длинную и сложную историю. Оксана при рождении весила почти на килограмм меньше, чем Наташа, и притом была синюшная и очень слабая, тогда как Наташа находилась в относительно благополучном состоянии. Это различие, обусловленное менее благоприятными обстоятельствами утробной жизни Оксаны, усилилось еще оттого, что в раннем детстве Оксана очень тяжело болела корью, а Наташа перенесла ее сравнительно легко. Оксана длительное время немного отставала в развитии и к 16 годам так и осталась несколько ниже Наташи ростом. Так как Наташа и родилась на 50 минут раньше Оксаны, то в связи с тем, что она опережала Оксану в развитии, ее стали дома считать «старшей», а Оксану — «младшей». Это внутрисемейное отношение между близнецами постепенно развивалось и крепло. В результате оно глубоко запечатлелось на всем «складе» высшей нервной деятельности близнецов, выразилось в том различии «характеров», которое выше отмечалось (Канаев, 1954).
Нечто подобное пришлось наблюдать на паре ОБ Ин. Одна из девочек этой пары — Нина — в раннем детстве имела воспаление уха, в связи с чем у нее возникла кривошея, сопровождавшаяся и некоторым отставанием в развитии по сравнению с ее близнячкой Тамарой. Последняя в этой паре оказалась в роли «ведущей», а Нина — «подчиненной», что вызвало целый ряд различий «склада» их высшей нервной деятельности (Канаев, 1937).
Как видно из этих первых попыток изучения физиологии высшей нервной деятельности, исследование ее в течение возможно длительного периода развития близнецов может дать ряд ценных сведении для понимания природы типа высшей нервной деятельности и ее «склада» в связи с условиями жизни, проливая новый свет на те материалы, которые получила психология по онтогенезу психики у близнецов (см. главу одиннадцатую).
Результатов исследования физиологии высшей нервной деятельности у соединенных близнецов (двух пар), изучавшихся в Москве, опубликовано еще очень мало (Анохин, 1939; Алексеева, 1941; Алексеева и Островская, 1953).
Физиология сна близнецов исследована еще мало. Павловская теория сна получила интересное подтверждение благодаря изучению двух пар соединенных близнецов Иры-Гали, родившихся в Москве в 1938 г. (Анохин, 1939; Алексеева, 1941). Многократно наблюдалось, что одна из близнячек спала, в то время как вторая находилась в состоянии бодрствования: смеялась, плакала и т. п. (рис. 53).
Кровеносная система этих близнецов сообщалась; следовательно, если бы сон зависел от содержания определенных веществ в крови (гуморальная теория сна), то близнецы засыпали бы всегда одновременно. Объяснение сна тормозным состоянием коры и нижележащих отделов мозга, как это делал И. П. Павлов, вполне соответствует наблюдавшейся на этой паре близнецов картине сна. Аналогичные данные получены и на второй паре соединенных близнецов (Алексеева и Островская, 1953). Интересно, что внутрипарные различия дыхания и частоты пульса этих близнецов, наблюдаемые во время их бодрствования, при одновременном глубоком сне в значительной мере исчезают.
Наблюдениями над сном целой серии пар ОБ и РБ установлено, что многие особенности сна — поза во время сна, глубина сна, его расстройства и т. д. — чаще конкордантны у ОБ, чем у РБ (Geyer, 1937). Однако такого рода исследование требует более точной постановки изучения физиологии сна, чем это делалось до сих нор, и привлечения эксперимента.
Как известно, во время сна моторная хронаксия у человека несколько увеличивается, т. е. возбудимость его тканей снижается в связи с сонным торможением, что обнаруживается в замедлении их реакции. Была сделана попытка, насколько мне известно, пока что единственная, исследовать моторную хронаксию у пары ОБ на первом году их жизни (Короткин и Крышова, 1948). Изучалась хронаксия флексоров и экстензоров пальцев руки. Обнаружено известное сходство динамики хронаксии у обоих близнецов, отличающее их от сверстников. Средние величины (из 100 отдельных измерений) хронаксии флексоров и экстензоров в бодрствующем и сонном состоянии оказались у близнецов очень схожими с таковыми у пары сверстников (рис. 158). К сожалению, это интересное исследование не было продолжено и развито.
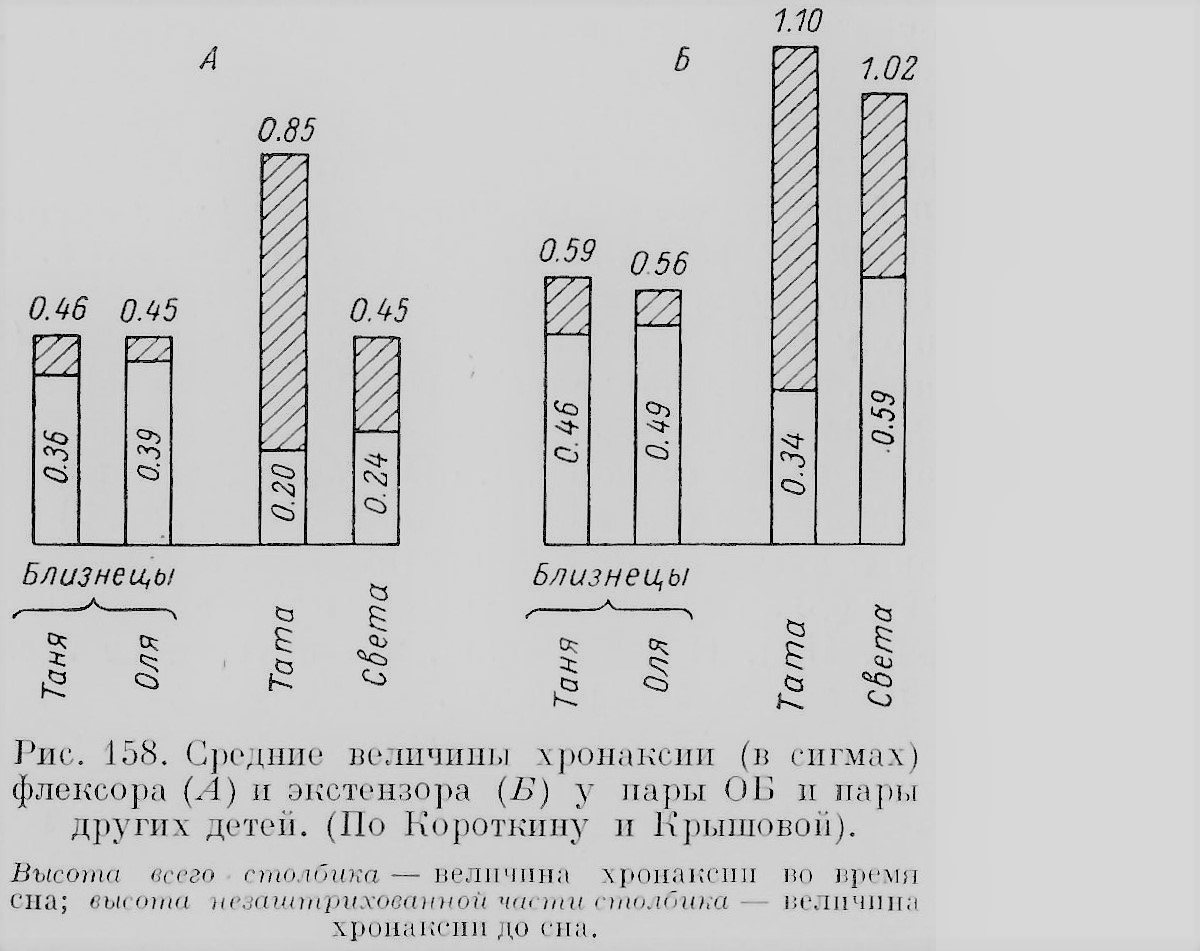
Глава одиннадцатая ПСИХОЛОГИЯ БЛИЗНЕЦОВ
Стремление измерить свойства психики человека, дать количественную оценку различным проявлениям ее, подобно тому как это делается в отношении различных физических особенностей человека антропометрией, — это стремление привело к созданию различных тестов, испытаний в действии психических свойств. Тесты — это серии вопросов или задач, в зависимости от числа и качества решений которых выводится количественная оценка испытуемой психической функции: «ума», «внимания», «темперамента», «фантазии», «памяти» и т. д. Так, например, для определения степени развития интеллекта употребляется система тестов Бине в различных редакциях. Для каждого возраста подобраны известные тесты, и по числу правильных ответов на них определяется «умственный возраст» испытуемого. Отношение этого «умственного возраста» к «паспортному возрасту» дает величину «коэффициента интеллигентности» (I. Q.), выражаемого обычно в процентах. Разумеется, тесты неизбежно носят условный характер: они рассчитаны на определенную шаблонность мышления субъектов испытуемой группы, и потому субъект с «необычным» складом мышления может получить низкую оценку. Очевидно, что и цифры, которыми измеряется «умственный возраст» и I. Q. имеют весьма относительную ценность, значительно меньшую, чем цифры, которыми определяются рост, вес и другие особенности тела испытуемого. Поэтому мы не будем рассматривать довольно обширную литературу о психике близнецов, изучаемой с помощью разных серий тестов, и ограничимся лишь одним примером изучения 50 пар ОБ и 50 РБ, у которых определялся I. Q. Прилагаемый график (рис. 159) дает итоговую картину. Мы видим, что внутрипарная разность группы ОБ меньше, чем группы РБ (Newman et. al., 1937).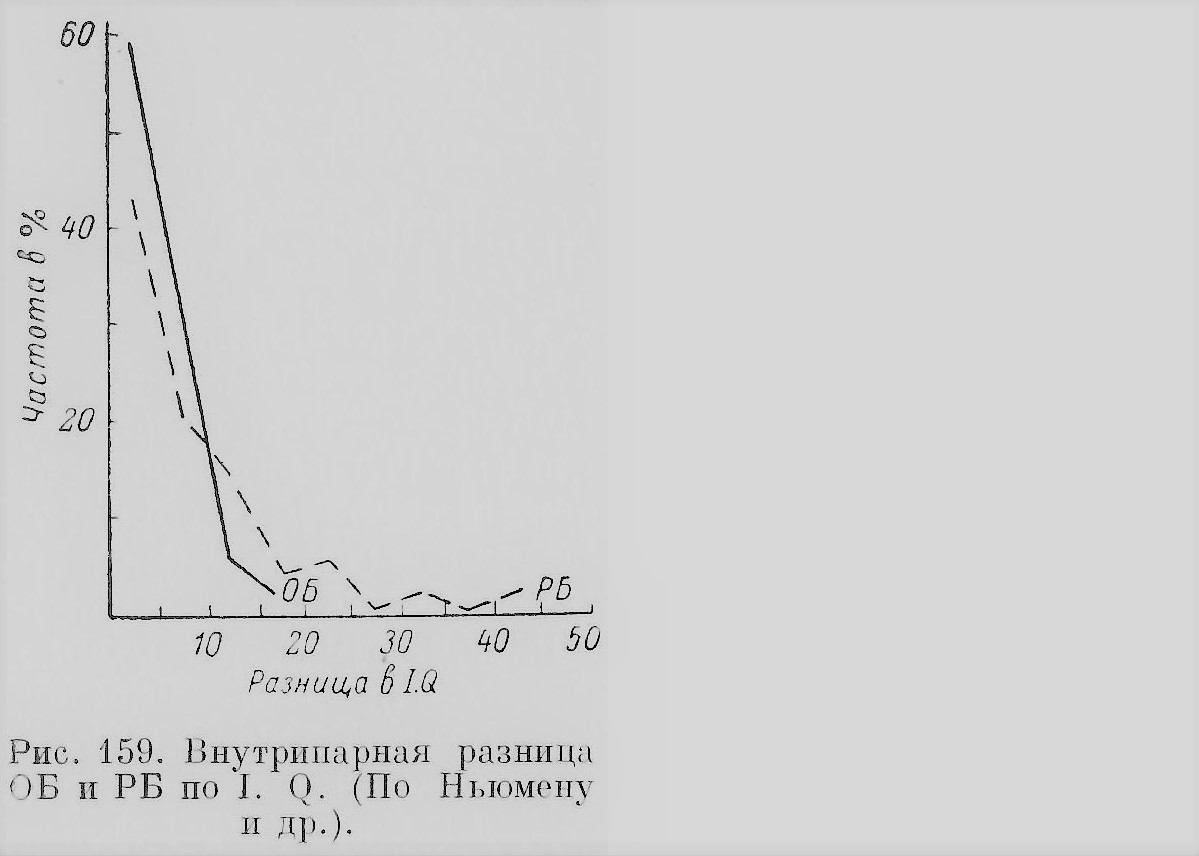 В той же работе авторы приходят к заключению, что в общем контраст внутрипарной разницы между ОБ и РБ меньше для психических свойств, чем для большинства физических, как например рост, промеры головы и т. п. Это наблюдение толкуется, как большая изменяемость факторами среды психических свойств, чем ряда физических (Newman, 1940b; Gedda, 1951, и др.). Попытки дифференцировать отдельные проявления психики в этом отношении с помощью разных серий тестов мало убедительны, но с известными ограничениями верным будет общее заключение о большей зависимости от среды психики, чем, например, роста. Это и понятно с точки зрения физиологии, так как мозг является важнейшим органом адаптации к окружающей действительности с ее постоянными изменениями.
Здесь еще нужно указать, что одно из последних исследований на близнецах с помощью тестов Роршаха (Schachter et Chatenet, 1953), произведенное на 23 парах ОБ и 7 парах РБ (детей и подростков), показало значительные внутрипарные различия психики ОБ, различие «личностей» ОБ одной пары. Хотя каждая из пар жила в общем в одинаковых условиях, всё же авторы считают, что внутрипарное психическое различие ОБ обусловлено факторами среды. Этот вывод находится в согласии с данными относительно различия физиологии высшей нервной деятельности у детей школьного возраста.
В 20-е и 30-е годы методом психологических тестов увлекались многие исследователи близнецов (ср. Lotze, 1937; Gedda, 1951); позже же некоторые стали предпочитать описательный метод (Smith, 1949; Verschuer, 1954, и др.).
К тестам по существу близко стоит, как прием, изучение почерка близнецов. Еще Гальтон обратил внимание на неразличимое сходство почерка одной пары близнецов. Рядом с этим он констатировал случаи различной степени внутрипарной разницы почерков. Это обнаружили и последующие исследователи. Известно, что при желании человек может коренным образом изменить свой почерк; имея, например, неразборчивый косой почерк, переделать его в четкий прямой и т. д. Не удивительно, что и близнецы одной пары могут, глядя по обстоятельствам, приобрести очень разные почерки, что не раз и наблюдалось. Делались многочисленные попытки сравнить почерки близнецов в разных аспектах (Gedda, 1951). Так, изучались четыре особенности почерков: «правильность» (степень математического сходства отдельных элементов), «равномерность» (ритм письма), «форма» (своеобразие начертания) и «угол наклона» букв (Lottig, 1931). ОБ и РБ по первому и четвертому признаку имели приблизительно одинаковую степень внутрипарного различия, тогда как по второму и третьему признаку ОБ были значительно более похожи внутрипарно, чем РБ. Также большее внутрипарное сходство обнаружилось у ОБ при изучении темпа письма — обычного, ускоренного и при состязании на скорость (Bracken, 1939а). По ряду свойств письма большее внутрипарное сходство ОБ установили и другие исследователи (Hermann, 1940: Roman-Goldzieher, 1945, и др.). Из 243 пар ОБ только у 5% было найдено полное внутрипарное сходство почерков. Интересно, что почерк пигопагов Хилтон не очень похож (одна из них пишет левой, другая — правой рукой), по все же имеет известное типическое для обеих сходство (Saudek a. Seaman, 1933). В итоге многочисленные работы над почерком близнецов дали пока что очень небольшой результат, что связано со сложностью самого процесса письма, малой изученностью физиологического механизма его и не всегда адекватной постановкой вопроса исследователями.
Неоднократно изучали и сравнивали школьную успеваемость близнецов по отметкам (Frischeisen-Kohler, 1930; Lehtovaara, 1938, и др.). Это, конечно, тоже своего рода тесты, далеко не совершенно отражающие способности, знания и труд школьников. Оказалось, что ОБ чаще имеют похожие отметки, чем РБ. Но точно судить по отметкам об одаренности детей, конечно, почти невозможно.
Более интересны данные относительно специальной одаренности близнецов, которую в некоторых случаях легче можно пронаблюдать и описать, чем такие менее заметные, нерезко выраженные психические свойства, как память или внимание. Так, например, сохранилось краткое описание близнецов-музыкантов Иоганна-Христофора и Иоганна-Амбросиуса Бахов. Последний был отцом знаменитого Иоганна-Себастиана Баха, сын которого, Филипп-Эммануил, так описал своего деда и его близнеца: «Они крайне любили друг друга. Они были так похожи, что даже их жены не могли их отличить. Они были чудом. . . для каждого, кто их видел. Речь, образ мысли — все у них было одинаково. В музыке также их нельзя было отличить. Они одинаково играли, одинаково развивали тему. Если один болел, то болел и другой. Они умерли вскоре один за другим». Очевидно, что эти близнецы были ОБ.
Сходство музыкальных способностей и их развития описано у пары ОБ — Вольфа и Билля Гейнц (Heinz), учеников Макса Регера, работавших дирижерами в двух разных театрах (рис. 160). Сходство их, не только внешнее, но и как музыкантов, было исключительно велико, даже Регер не мог их различить. Рассказывают такой случай: оба готовили одну и ту же оперу, каждый в своем театре. Сходство их интерпретации и манеры дирижировать было так велико, что один из них мог заменить другого после антракта при исполнении оперы, и никто из исполнителей (певцов или оркестрантов), а также из публики, не замечал, что дирижирует другой человек (Verschuer, 1930).
Однако сходство музыкальных способностей пары ОБ не всегда бывает так полно, как у вышеописанных близнецов. Мне известна пара ОБ, специалисток по органу, которые, по свидетельству их учителя, проф. И. А. Браудо, чрезвычайно похожи по разным признакам музыкального дарования (слух, музыкальная память, исполнительские данные и т. д.), но различаются в смысле интерпретации одной и той же вещи при ее исполнении, т. е. каждая играет ее по-своему. При этом близнецы жили до взрослого возраста вместе, вместе учились в консерватории и кончили ее под руководством одного и того же профессора. Внешне обе очень похожи, по по выражению глаз их можно легко различить; здесь сказывается различие склада высшей нервной деятельности, которое обнаруживается в их игре.
В области других способностей можно указать на близнецов-ученых Огюста и Жана Пиккар (рис. 160), внешне очень похожих. Оба имели сходные способности и оба увлекались естествознанием. Огюст стал физиком, Жан — химиком. Первый получил кафедру в Бельгии, второй — в Америке. Оба известны полетом в стратосферу (Lotze, 1937). Так сходство дарований при сходстве обучения и благоприятных социальных условий переходит дальше в сходство профессии и связанного с ней склада жизни и жизненной карьеры.
В связи с затронутыми здесь вопросами надо также коснуться проблемы психической дефективности близнецов, не раз обсуждавшейся в литературе (ср. Miettinen, 1954). Указывалось, что идиоты, олигофреники, психически больные и т. д. чаще встречаются среди близнецов, чем среди массы населения (Rosanoff a. Inman-Kane, 1934, и др.). Объясняется это тем, что близнецы часто бывают недоносками; кроме того, трудности утробной жизни и родов также могут сказываться отрицательно (Brander, 1937, и др.). По некоторым данным, близнецы не дают большего процента умственно отсталых (2.8(+/-)1.43), чем недоноски-одиночки (3.5(+/-)0.8), однако больший процент, чем доношенные дети (0.7(+/-)0.3) (Alm, 1953). Но есть исследователи, которые вообще отрицают какое-либо существенное различие в физическом и умственном развитии между недоносками и во время родившимися младенцами (Hess, Mohr а. Bartelme, 1934; Crosse, 1952, и др.). Таким образом, весь этот вопрос нуждается в дальнейшем изучении. Он еще менее разработан в отношении «сверхблизнецов», т. е. троен, четверен и т. д. Некоторое увеличение процента умственно дефективных у этого рода близнецов, по-видимому, имеется, как например это установлено на финском материале, где среди 445 детей-троен оказалось 7.0(+/-)1.2% умственно неполноценных (Miettinen, 1954).
Одно из важнейших преимуществ близнецов было использовано в экспериментальной психологии — это внутрипарное сходство ОБ. Один из близнецов служил для опыта, другой — для контроля. Это метод «взаимоконтроля» близнецов («cotwin control»), введенный Гизеллом (Gesell, 1929—1952). Разумеется, что не всякая пара ОБ может служить для подобного эксперимента, а лишь такая, партнеры которой в достаточной мере похожи. В опытах с моторикой использовалась также киносъемка для сравнения различных звеньев одного и разных экспериментов. Так, в возрасте 46 недель один из ОБ — экспериментальный — тренировался в лазании по лестнице в 5 ступеней и после 52 недель влезал на нее за 26 секунд. Контрольный в возрасте 53 недель за 2 недели тренировки не только догнал, но даже перегнал своего партнера. Эти и дальнейшие данные дают материал для суждения о влиянии возраста на тренировку. Аналогичное исследование на той же паре велось с развитием речи (Strayer, 1930). Детей разлучили и экспериментального близнеца с 84-й по 89-ю неделю тренировали в развитии речи. С 89-й недели в течение 4 недель тренировали контрольного близнеца. Последний достиг большего успеха через 28 дней, чем первый за 35 дней. В течение 3 месяцев разница стерлась (подробнее об этой паре см. главу тринадцатую «Длительное изучение развития одной пары ОБ»).
Метод взаимоконтроля близнецов был развит советскими психологами (Миренова, 1932, 1934; Миренова и Колбановский, 1934; Лурия и Миренова, 1936б, и др.). В одной из серий исследований изучалась конструктивная деятельность близнецов, которым предлагалось построить из кубиков различные фигуры (зданий и т. п.) двумя разными методами: простым копированием уже готового сооружения из кубиков (Э — элементарный метод) или постройкой из кубиков по модели (М — метод моделей), оклеенной бумагой, что не позволяло пассивно копировать ее, а требовало известных конструктивных усилий. В опыте участвовало 5 пар ОБ в возрасте 51/2—6 лет. Из каждой пары один близнец тренировался пассивным методом (Э), другой — активным (М). Второй метод способствовал заметному развитию не только конструктивной деятельности ребенка, но и сопровождающей ее более глубокой перестройки психики ребенка — его «восприятия» окружающего, речи и т. д. Эффект от такой тренировки, лежащий за пределами самого конструктивного навыка с кубиками, продолжался свыше года (Лурия и Миренова, 1936б). Интересно было бы произвести анализ этих явлений с целью выяснения условно-рефлекторных механизмов, управляющих ими.
По-видимому, метод взаимоконтроля близнецов может быть применен для решения различных вопросов воспитания и обучения в дошкольном и школьном возрасте с целью проверки эффективности различных педагогических приемов и методов. Недавно с помощью этого метода были получены интересные данные по развитию речи у пары ОБ (Лурия и Юдович, 1956), о которых сказано подробнее в связи с изучением речи у близнецов (см. далее).
Метод взаимоконтроля нашел также применение в клинике (Gesell, 1936; Босик и Пасынков, 1936; Greene, 1942; Mundy, 1953, и др.), но пока еще недостаточно.
Здесь мы касаемся, и уже не первый раз, одного общего вопроса, очень важного для изучения близнецов, — отношения близнецов к окружающей среде. Природное сходство пары ОБ неизбежно приводит к тому, что они стремятся к похожим условиям среды, к деятельности, соответствующей их природным данным, и т. д. Так, например, если пара ОБ будет со слабой нервной системой, то оба будут тяготеть к обстановке, достаточно тихой и уединенной, в которой их нервная система не будет перегружена от воздействия внешних раздражений. Наоборот, природное различие РБ обнаружится легко в том, что «нравиться» им будет разная обстановка, разная деятельность и т. д. Таким образом, из тех возможностей, которые представляет одна и та же среда, в ряде случаев пара ОБ будет выбирать условия более похожие, чем пара РБ (Lehtovaara, 1938, и др.). Этот момент — выбор условий в соответствии с природными данными — является одной из существенных сторон в проблеме наследственность—среда, говорящий об их тесной взаимозависимости (ср. Zazzo, 1955). Такой «выбор» условий незаметно переходит в «создание» условий, соответствующих потребности данного индивидуума. В биографии человека, в частности биографиях пары близнецов, можно высмотреть проявление этой сложной диалектики взаимоотношения природы человека и среды, в которой он живет. Это особенно заметно, конечно, в случаях ярко одаренных близнецов, когда талант может играть значительную роль во всем складе жизни человека, как в упомянутых выше случаях, касающихся музыкантов или ученых.
Любопытно было бы глубже изучить различие среды, при кажущемся внешнем сходстве ее, для четверни, например, состоящей из пары ОБ и пары РБ: что в окружающей их среде будут «выбирать» такие вместе растущие дети? К сожалению, такие исследования систематически еще не велись. Очень важным элементом среды для всякого близнеца является его партнер, другой близнец. К этому вопросу мы теперь и перейдем.
Многие исследователи интересовались взаимоотношением близнецов (Bracken, 1936, 1939b; Schiller, 1937; Lehtovaara, 1938; Smith, 1949; Gedda, 1951; Verschuer, 1954, и др.). Пара близнецов — это своеобразный маленький коллектив. В силу большего внутрипарного сходства связи между парой ОБ, по-видимому, часто более интимны и глубоки, чем между парой РБ. С возрастом, с возникновением разнообразных новых связей взаимоотношения между партнерами обычно ослабевают. Считается, что у близнецов мужского пола это психическое расхождение происходит легче, чем у близнецов женского пола (Lehtovaara, 1938; Smith, 1949).
Характер взаимоотношений у разных пар очень разнообразен. Собственно, каждая пара близнецов имеет свои особенности, является единственной в своем роде, как вообще неповторима индивидуальность отдельных людей. Если попытаться схематизировать внутрипарные отношения близнецов, то можно сказать, что одной из любопытных особенностей является то, что можно было бы условно назвать «первенством». Дело в том, что нередко складываются такие отношения, постепенно все больше дифференцирующиеся, когда один из близнецов занимает место «ведущего» («лидера»), а другой соответственно «ведомого», иначе говоря, «главенствующего» и «подчиненного», о чем уже попутно речь была выше. От отношений полного, так сказать, «равноправия», когда ни один из двух не является «лидером», через разнообразные формы слабого и временного проявления признаков главенства у одного из близнецов, иногда с переходом этой функции попеременно от одного к другому, дело доходит до полного и постоянного подчинения одного близнеца другому с соответствующим распределением ролей в труде, играх и т. д. Получается своего рода «поляризация» во взаимоотношениях внутри пары близнецов, но именно такая «поляризация» и свидетельствует о том, что эта пара образует своеобразное единое целое (ср. Lehtovaara, 1938). Конечно, разные типы таких отношений — от «равноправия» до «порабощения» — далеко не всегда развиваются одним путем и не всегда доходят до крайнего предела. Очень часто они достигают только определенных степеней по этой линии развития взаимоотношений и дальше не идут. По-видимому, разные ступени такого рода вполне совместимы с разной степенью любви и дружбы между членами одной пары. Вражда и ненависть между близнецами, по аналогии с отношениями между Ромулом и Ремом, по-видимому, очень редко встречаются (Verschuer, 1954).
В той же работе авторы приходят к заключению, что в общем контраст внутрипарной разницы между ОБ и РБ меньше для психических свойств, чем для большинства физических, как например рост, промеры головы и т. п. Это наблюдение толкуется, как большая изменяемость факторами среды психических свойств, чем ряда физических (Newman, 1940b; Gedda, 1951, и др.). Попытки дифференцировать отдельные проявления психики в этом отношении с помощью разных серий тестов мало убедительны, но с известными ограничениями верным будет общее заключение о большей зависимости от среды психики, чем, например, роста. Это и понятно с точки зрения физиологии, так как мозг является важнейшим органом адаптации к окружающей действительности с ее постоянными изменениями.
Здесь еще нужно указать, что одно из последних исследований на близнецах с помощью тестов Роршаха (Schachter et Chatenet, 1953), произведенное на 23 парах ОБ и 7 парах РБ (детей и подростков), показало значительные внутрипарные различия психики ОБ, различие «личностей» ОБ одной пары. Хотя каждая из пар жила в общем в одинаковых условиях, всё же авторы считают, что внутрипарное психическое различие ОБ обусловлено факторами среды. Этот вывод находится в согласии с данными относительно различия физиологии высшей нервной деятельности у детей школьного возраста.
В 20-е и 30-е годы методом психологических тестов увлекались многие исследователи близнецов (ср. Lotze, 1937; Gedda, 1951); позже же некоторые стали предпочитать описательный метод (Smith, 1949; Verschuer, 1954, и др.).
К тестам по существу близко стоит, как прием, изучение почерка близнецов. Еще Гальтон обратил внимание на неразличимое сходство почерка одной пары близнецов. Рядом с этим он констатировал случаи различной степени внутрипарной разницы почерков. Это обнаружили и последующие исследователи. Известно, что при желании человек может коренным образом изменить свой почерк; имея, например, неразборчивый косой почерк, переделать его в четкий прямой и т. д. Не удивительно, что и близнецы одной пары могут, глядя по обстоятельствам, приобрести очень разные почерки, что не раз и наблюдалось. Делались многочисленные попытки сравнить почерки близнецов в разных аспектах (Gedda, 1951). Так, изучались четыре особенности почерков: «правильность» (степень математического сходства отдельных элементов), «равномерность» (ритм письма), «форма» (своеобразие начертания) и «угол наклона» букв (Lottig, 1931). ОБ и РБ по первому и четвертому признаку имели приблизительно одинаковую степень внутрипарного различия, тогда как по второму и третьему признаку ОБ были значительно более похожи внутрипарно, чем РБ. Также большее внутрипарное сходство обнаружилось у ОБ при изучении темпа письма — обычного, ускоренного и при состязании на скорость (Bracken, 1939а). По ряду свойств письма большее внутрипарное сходство ОБ установили и другие исследователи (Hermann, 1940: Roman-Goldzieher, 1945, и др.). Из 243 пар ОБ только у 5% было найдено полное внутрипарное сходство почерков. Интересно, что почерк пигопагов Хилтон не очень похож (одна из них пишет левой, другая — правой рукой), по все же имеет известное типическое для обеих сходство (Saudek a. Seaman, 1933). В итоге многочисленные работы над почерком близнецов дали пока что очень небольшой результат, что связано со сложностью самого процесса письма, малой изученностью физиологического механизма его и не всегда адекватной постановкой вопроса исследователями.
Неоднократно изучали и сравнивали школьную успеваемость близнецов по отметкам (Frischeisen-Kohler, 1930; Lehtovaara, 1938, и др.). Это, конечно, тоже своего рода тесты, далеко не совершенно отражающие способности, знания и труд школьников. Оказалось, что ОБ чаще имеют похожие отметки, чем РБ. Но точно судить по отметкам об одаренности детей, конечно, почти невозможно.
Более интересны данные относительно специальной одаренности близнецов, которую в некоторых случаях легче можно пронаблюдать и описать, чем такие менее заметные, нерезко выраженные психические свойства, как память или внимание. Так, например, сохранилось краткое описание близнецов-музыкантов Иоганна-Христофора и Иоганна-Амбросиуса Бахов. Последний был отцом знаменитого Иоганна-Себастиана Баха, сын которого, Филипп-Эммануил, так описал своего деда и его близнеца: «Они крайне любили друг друга. Они были так похожи, что даже их жены не могли их отличить. Они были чудом. . . для каждого, кто их видел. Речь, образ мысли — все у них было одинаково. В музыке также их нельзя было отличить. Они одинаково играли, одинаково развивали тему. Если один болел, то болел и другой. Они умерли вскоре один за другим». Очевидно, что эти близнецы были ОБ.
Сходство музыкальных способностей и их развития описано у пары ОБ — Вольфа и Билля Гейнц (Heinz), учеников Макса Регера, работавших дирижерами в двух разных театрах (рис. 160). Сходство их, не только внешнее, но и как музыкантов, было исключительно велико, даже Регер не мог их различить. Рассказывают такой случай: оба готовили одну и ту же оперу, каждый в своем театре. Сходство их интерпретации и манеры дирижировать было так велико, что один из них мог заменить другого после антракта при исполнении оперы, и никто из исполнителей (певцов или оркестрантов), а также из публики, не замечал, что дирижирует другой человек (Verschuer, 1930).
Однако сходство музыкальных способностей пары ОБ не всегда бывает так полно, как у вышеописанных близнецов. Мне известна пара ОБ, специалисток по органу, которые, по свидетельству их учителя, проф. И. А. Браудо, чрезвычайно похожи по разным признакам музыкального дарования (слух, музыкальная память, исполнительские данные и т. д.), но различаются в смысле интерпретации одной и той же вещи при ее исполнении, т. е. каждая играет ее по-своему. При этом близнецы жили до взрослого возраста вместе, вместе учились в консерватории и кончили ее под руководством одного и того же профессора. Внешне обе очень похожи, по по выражению глаз их можно легко различить; здесь сказывается различие склада высшей нервной деятельности, которое обнаруживается в их игре.
В области других способностей можно указать на близнецов-ученых Огюста и Жана Пиккар (рис. 160), внешне очень похожих. Оба имели сходные способности и оба увлекались естествознанием. Огюст стал физиком, Жан — химиком. Первый получил кафедру в Бельгии, второй — в Америке. Оба известны полетом в стратосферу (Lotze, 1937). Так сходство дарований при сходстве обучения и благоприятных социальных условий переходит дальше в сходство профессии и связанного с ней склада жизни и жизненной карьеры.
В связи с затронутыми здесь вопросами надо также коснуться проблемы психической дефективности близнецов, не раз обсуждавшейся в литературе (ср. Miettinen, 1954). Указывалось, что идиоты, олигофреники, психически больные и т. д. чаще встречаются среди близнецов, чем среди массы населения (Rosanoff a. Inman-Kane, 1934, и др.). Объясняется это тем, что близнецы часто бывают недоносками; кроме того, трудности утробной жизни и родов также могут сказываться отрицательно (Brander, 1937, и др.). По некоторым данным, близнецы не дают большего процента умственно отсталых (2.8(+/-)1.43), чем недоноски-одиночки (3.5(+/-)0.8), однако больший процент, чем доношенные дети (0.7(+/-)0.3) (Alm, 1953). Но есть исследователи, которые вообще отрицают какое-либо существенное различие в физическом и умственном развитии между недоносками и во время родившимися младенцами (Hess, Mohr а. Bartelme, 1934; Crosse, 1952, и др.). Таким образом, весь этот вопрос нуждается в дальнейшем изучении. Он еще менее разработан в отношении «сверхблизнецов», т. е. троен, четверен и т. д. Некоторое увеличение процента умственно дефективных у этого рода близнецов, по-видимому, имеется, как например это установлено на финском материале, где среди 445 детей-троен оказалось 7.0(+/-)1.2% умственно неполноценных (Miettinen, 1954).
Одно из важнейших преимуществ близнецов было использовано в экспериментальной психологии — это внутрипарное сходство ОБ. Один из близнецов служил для опыта, другой — для контроля. Это метод «взаимоконтроля» близнецов («cotwin control»), введенный Гизеллом (Gesell, 1929—1952). Разумеется, что не всякая пара ОБ может служить для подобного эксперимента, а лишь такая, партнеры которой в достаточной мере похожи. В опытах с моторикой использовалась также киносъемка для сравнения различных звеньев одного и разных экспериментов. Так, в возрасте 46 недель один из ОБ — экспериментальный — тренировался в лазании по лестнице в 5 ступеней и после 52 недель влезал на нее за 26 секунд. Контрольный в возрасте 53 недель за 2 недели тренировки не только догнал, но даже перегнал своего партнера. Эти и дальнейшие данные дают материал для суждения о влиянии возраста на тренировку. Аналогичное исследование на той же паре велось с развитием речи (Strayer, 1930). Детей разлучили и экспериментального близнеца с 84-й по 89-ю неделю тренировали в развитии речи. С 89-й недели в течение 4 недель тренировали контрольного близнеца. Последний достиг большего успеха через 28 дней, чем первый за 35 дней. В течение 3 месяцев разница стерлась (подробнее об этой паре см. главу тринадцатую «Длительное изучение развития одной пары ОБ»).
Метод взаимоконтроля близнецов был развит советскими психологами (Миренова, 1932, 1934; Миренова и Колбановский, 1934; Лурия и Миренова, 1936б, и др.). В одной из серий исследований изучалась конструктивная деятельность близнецов, которым предлагалось построить из кубиков различные фигуры (зданий и т. п.) двумя разными методами: простым копированием уже готового сооружения из кубиков (Э — элементарный метод) или постройкой из кубиков по модели (М — метод моделей), оклеенной бумагой, что не позволяло пассивно копировать ее, а требовало известных конструктивных усилий. В опыте участвовало 5 пар ОБ в возрасте 51/2—6 лет. Из каждой пары один близнец тренировался пассивным методом (Э), другой — активным (М). Второй метод способствовал заметному развитию не только конструктивной деятельности ребенка, но и сопровождающей ее более глубокой перестройки психики ребенка — его «восприятия» окружающего, речи и т. д. Эффект от такой тренировки, лежащий за пределами самого конструктивного навыка с кубиками, продолжался свыше года (Лурия и Миренова, 1936б). Интересно было бы произвести анализ этих явлений с целью выяснения условно-рефлекторных механизмов, управляющих ими.
По-видимому, метод взаимоконтроля близнецов может быть применен для решения различных вопросов воспитания и обучения в дошкольном и школьном возрасте с целью проверки эффективности различных педагогических приемов и методов. Недавно с помощью этого метода были получены интересные данные по развитию речи у пары ОБ (Лурия и Юдович, 1956), о которых сказано подробнее в связи с изучением речи у близнецов (см. далее).
Метод взаимоконтроля нашел также применение в клинике (Gesell, 1936; Босик и Пасынков, 1936; Greene, 1942; Mundy, 1953, и др.), но пока еще недостаточно.
Здесь мы касаемся, и уже не первый раз, одного общего вопроса, очень важного для изучения близнецов, — отношения близнецов к окружающей среде. Природное сходство пары ОБ неизбежно приводит к тому, что они стремятся к похожим условиям среды, к деятельности, соответствующей их природным данным, и т. д. Так, например, если пара ОБ будет со слабой нервной системой, то оба будут тяготеть к обстановке, достаточно тихой и уединенной, в которой их нервная система не будет перегружена от воздействия внешних раздражений. Наоборот, природное различие РБ обнаружится легко в том, что «нравиться» им будет разная обстановка, разная деятельность и т. д. Таким образом, из тех возможностей, которые представляет одна и та же среда, в ряде случаев пара ОБ будет выбирать условия более похожие, чем пара РБ (Lehtovaara, 1938, и др.). Этот момент — выбор условий в соответствии с природными данными — является одной из существенных сторон в проблеме наследственность—среда, говорящий об их тесной взаимозависимости (ср. Zazzo, 1955). Такой «выбор» условий незаметно переходит в «создание» условий, соответствующих потребности данного индивидуума. В биографии человека, в частности биографиях пары близнецов, можно высмотреть проявление этой сложной диалектики взаимоотношения природы человека и среды, в которой он живет. Это особенно заметно, конечно, в случаях ярко одаренных близнецов, когда талант может играть значительную роль во всем складе жизни человека, как в упомянутых выше случаях, касающихся музыкантов или ученых.
Любопытно было бы глубже изучить различие среды, при кажущемся внешнем сходстве ее, для четверни, например, состоящей из пары ОБ и пары РБ: что в окружающей их среде будут «выбирать» такие вместе растущие дети? К сожалению, такие исследования систематически еще не велись. Очень важным элементом среды для всякого близнеца является его партнер, другой близнец. К этому вопросу мы теперь и перейдем.
Многие исследователи интересовались взаимоотношением близнецов (Bracken, 1936, 1939b; Schiller, 1937; Lehtovaara, 1938; Smith, 1949; Gedda, 1951; Verschuer, 1954, и др.). Пара близнецов — это своеобразный маленький коллектив. В силу большего внутрипарного сходства связи между парой ОБ, по-видимому, часто более интимны и глубоки, чем между парой РБ. С возрастом, с возникновением разнообразных новых связей взаимоотношения между партнерами обычно ослабевают. Считается, что у близнецов мужского пола это психическое расхождение происходит легче, чем у близнецов женского пола (Lehtovaara, 1938; Smith, 1949).
Характер взаимоотношений у разных пар очень разнообразен. Собственно, каждая пара близнецов имеет свои особенности, является единственной в своем роде, как вообще неповторима индивидуальность отдельных людей. Если попытаться схематизировать внутрипарные отношения близнецов, то можно сказать, что одной из любопытных особенностей является то, что можно было бы условно назвать «первенством». Дело в том, что нередко складываются такие отношения, постепенно все больше дифференцирующиеся, когда один из близнецов занимает место «ведущего» («лидера»), а другой соответственно «ведомого», иначе говоря, «главенствующего» и «подчиненного», о чем уже попутно речь была выше. От отношений полного, так сказать, «равноправия», когда ни один из двух не является «лидером», через разнообразные формы слабого и временного проявления признаков главенства у одного из близнецов, иногда с переходом этой функции попеременно от одного к другому, дело доходит до полного и постоянного подчинения одного близнеца другому с соответствующим распределением ролей в труде, играх и т. д. Получается своего рода «поляризация» во взаимоотношениях внутри пары близнецов, но именно такая «поляризация» и свидетельствует о том, что эта пара образует своеобразное единое целое (ср. Lehtovaara, 1938). Конечно, разные типы таких отношений — от «равноправия» до «порабощения» — далеко не всегда развиваются одним путем и не всегда доходят до крайнего предела. Очень часто они достигают только определенных степеней по этой линии развития взаимоотношений и дальше не идут. По-видимому, разные ступени такого рода вполне совместимы с разной степенью любви и дружбы между членами одной пары. Вражда и ненависть между близнецами, по аналогии с отношениями между Ромулом и Ремом, по-видимому, очень редко встречаются (Verschuer, 1954).
 В качестве примера резко выраженной «поляризации» взаимоотношений пары ОБ при очень большой взаимной дружбе рассмотрим Наташу и Ноэми (Эму) Ботв., 10 лет (рис. 161). Они родились с очень большим интервалом, почти в 5 дней (имели, по-видимому, разные плаценты и оболочки); с первых месяцев стали проявлять заметное различие характеров: Наташа была более активной по сравнению с Эмой. В раннем детстве они были очень дружны. Как нередко бывает в подобных случаях, их любимым местоимением было «мы», которым они, впрочем, злоупотребляли в некоторых случаях, например, заявив однажды родителям: «Мы сегодня ночью видели во сне. . .», и далее, перебивая друг друга, стали рассказывать содержание сна. Дружба детей не пострадала от того, что более деятельная Наташа стала постепенно инициатором в организации игр, в выполнении различных домашних дел и поручений, представителем близнецов во внешнем мире и т. д., а Эма — ее подсобницей, совершенно пассивной, предоставляющей Наташе активную роль. Это взаимоотношение очень наглядно выступило в одном мелком эпизоде. Эма, во время разговора в лаборатории после опыта, вертела гайку на одном приборе, гайка отлетела и покатилась на пол. Я попросил найти эту гайку. Наташа сразу бросилась искать ее под столом и под соседним шкафом, а Эма, заложив руки за спину спокойно наблюдала за близнячкой. На мой вопрос, почему же ищет Наташа, а не она, отвинтившая гайку. Эма ответила: «Такие дела у нас делает Наташа». При подобном взаимоотношении Эма охотно и покорно подчиняется инициативе Наташи, пользуется ее активностью, а Наташа с удовольствием «командует» и сознает себя незаменимой в их близнецовом коллективе. Степень, которой достигает такого рода «поляризация» у этих близнецов, вредно отзывается на обеих, развивая своеобразную односторонность у каждой из девочек. В таких случаях, как этот, очень ясно выступает значение одного близнеца для другого, как очень действенного и постоянно влияющего фактора среды. С возрастом, с развитием детей такие отношения все больше дифференцируются, пока не достигнут известного предела, также, вероятно, связанного с известной стадией онтогенеза.
Те взаимоотношения, которые так разнообразно слагаются в пределах каждой пары близнецов, особенно любопытно развертываются в близнецовых коллективах с большим числом членов: в тройнях, четвернях и пятернях. Мы рассмотрим два примера: однояйцевую четверню Морлок (Gardner a. Newman, 1943) и однояйцевую пятерню Дионн (Schwesinger, 1940, и др.).
Девочки Морлок (рис. 129) в возрасте 10 лет имели известную разницу в весе и росте. Интересно, что у этих близнецов между величиной роста и веса, с одной стороны, и степенью умственного развития (I.Q.),c другой, была известная корреляция, так же как и у Дионн. Это интересное явление, пока еще недостаточно изученное, могло послужить основой внутренней организации коллектива данной четверни. Заправилой, «лидером» всей группы стала самая крупная близнячка, опережавшая остальных в развитии. Она была представителем интересов четверни в окружающем мире, «депутатом». Самая маленькая в коллективе была «бэби», которую опекали остальные. Две другие взяли на себя роли своего рода затейников: одна была «клоун», задира и шутник, а другая — «артист» в силу большей склонности к рисованию и музыке. Конечно, это не были очень уж прочно фиксированные «роли», но все же они свидетельствовали о внутренней организации коллектива и известном распределении функций между его членами, известной «специализации» их. Морлок пели и танцевали как дружный ансамбль. Интересно, что, в отличие от Дионн, изолированных от внешнего мира, Морлок жили, как прочие дети, в обычном общении со своими сверстниками в школе и вообще с окружающим миром. По умственному развитию они были несколько выше среднего.
У пятерни Дионн (рис. 131), речь о которых подробнее идет в другом месте (глава девятая), внутри их коллектива сложились до известной степени аналогичные отношения: у них также был «лидер» коллектива, была самая «младшая» — «бэби» (Мари), и остальные трое уже с 4 лет проявляли известные индивидуальные особенности. К сожалению, мне неизвестна ни одна работа, в которой бы описывалось развитие индивидуальных особенностей каждой из пяти девочек с раннего возраста (Дионн родились в 1934 г.) в системе их маленького мирка как замкнутого целого.
Одна из попыток проникнуть во внутреннюю структуру коллектива пятерни Дионн принадлежит экологу Элли (Аllее, 1951). В течение 14 месяцев, с 22-го по 36-й месяц жизни детей, наблюдались «контакты» детей друг с другом и велся счет таких соприкосновений. При этом из общего числа различных «контактов» выделялись те из них, когда одни ребенок по своей инициативе адресовался к другому и когда ребенок отвечал на обращение к нему. Отдельные дети из пятерни Дионн заметно отличаются друг от друга в этом отношении, и различия в некоторых случаях носят стойкий и длительный характер. Так, Анетта все время занимала первое место, Цецилия — всегда второе. Остальные имели не столь постоянные места. Между числом контактов и интеллектуальным уровнем (по тестам) корреляции не наблюдалось. Так, самая развитая из группы — Ивонна — по инициативе к контактам занимает последнее место, а «бэби» группы — Мари — среднее место и т. д. Конечно, эти данные Элли лишь очень схематично, односторонне, по аналогии с его опытами с курами, и скудно освещают интересный вопрос структуры такого коллектива и причины этой структуры не раскрывают. Методически изучение этого вопроса должным образом вообще еще не поставлено.
Как одно из проявлений взаимоотношения близнецов надо, по-видимому, рассматривать вопрос об одежде близнецов, хотя он содержит, конечно, и другие элементы. Опрос и наблюдения показали, что пары ОБ чаще, чем пары РБ, желают иметь одинаковую одежду. В этом, очевидно, обнаруживается их чувство солидарности, симпатии друг к другу. Вероятно, что в ряде случаев РБ хотят различием одежды подчеркнуть свою независимость друг от друга, нежелание слыть за близнецов и т. п. (Lehtovaara, 1938; Gedda, 1951, и др.). Однако, как отмечает Гедда, желание иметь одинаковую одежду не всегда совпадает с желанием надевать одежду своего близнеца. Здесь выступает сознание своей индивидуальности, несмотря на симпатию и преданность своему близнецу.
Тот коллектив, который образуют близнецы, особенно если он состоит из большего числа детей, чем двое, иногда приобретает тенденцию замыкаться от окружающего мира, особенно, если близнецы вырабатывают своеобразную речь, понятную только им. Это вредно отражается на развитии близнецов, они отстают от сверстников в смысле психического развития и т. д. (Zazzo, 1952, и др.).
Близнецовый коллектив входит обычно в состав семьи как более крупного коллектива. Влияние отца и матери, братьев и сестер на развитие близнецов, влияние разницы в отношении к близнецам других членов семьи на близнецов и их взаимоотношения — вопросы очень важные для оценки роли социальной среды на формирование психики близнецов. Например, одна из девочек пары ОБ, Нина Ин. о которой речь идет побробнее в другом месте, страдала кривошеей; отец явно больше любил ее нормальную близнячку. Этим, конечно, усиливалось сознание неполноценности у пострадавшей с вытекающими отсюда последствиями. Многообразное и очень важное влияние семьи на развитие психики близнецов еще очень мало исследовано. Примером своеобразного жизненного эксперимента, показывающего важность роли семейной среды, является случай с парой ОБ, описанный Швезингер (Schwesinger, 1952). ОБ Эльвира и Эсфирь росли в разных семьях: первая в семье своей матери, вышедшей вторично замуж и имевшей еще других детей, с которыми Эльвира дружно жила; вторая у суровой приемной матери, в одиночестве и трудных условиях. В материальном отношении обе близнячки жили примерно на одном уровне достаточности. Жизнь Эльвиры сложилась сравнительно удачно, тогда как Эсфирь, замкнутая, неуравновешенная, стала морфинисткой и рано погибла, по-видимому, покончив самоубийством.
Были попытки выяснить социальные отношения близнецов вне семьи — в школьной среде (Lehtovaara, 1938). Это трудная задача, и пока что ее нельзя считать решенной. Выбор детей, с которыми близнецы дружат, совместно играют и т. д., их возраст, пол, число их и проч. — все это учитывалось. Оказалось, что выбор ОБ в отношении товарищей в том же школьном коллективе в общем более похож, чем РБ. Это один из примеров того избирательного отношения к среде, о котором речь была выше.
Социальная ориентация близнецов обнаруживается также в их детских планах относительно будущей профессии. Около 75% пар ОБ имело похожие планы, тогда как только около 50% пар РБ высказало в той или иной мере схожие проекты своего будущего (Lehtovaara, 1938).
Все вышеприведенные данные об отношении близнецов к их социальной среде теснейшим образом связаны с вопросами способов выражения, самообнаружения, общения, т. е. речи, рисунка, мимики, жестикуляции и т. д. К этой группе вопросов мы теперь и переходим.
О развитии речи у близнецов говорится в другом месте (см. главу тринадцатую). С психологических позиций речь близнецов еще очень мало изучена, и физиология речи у близнецов по существу вовсе не изучена. Попытка анализа речи 12 пар близнецов обоих типов по определенной системе (Шейдта), различающей 5 основных классов слов, не обнаружила существенно большего внутрипарного сходства ОБ по сравнению с РБ (Gottschick, 1939). Далее были сделаны попытки установить связь особенностей речи с характеристикой «личности» близнецов и конституциональными типами, моторикой, «личным темпом» и т. д. (Luchsinger, 1940). Все эти попытки малоубедительны с методической и методологической стороны. Например, изучение «внутренней речи» (мыслей и представлений без произнесения слов испытуемым) велось интроспективным методом, т. е. на основании словесного отчета испытуемого после выполнения им определенных заданий (например, «продумать» без произнесения слов 7 пунктов из таблицы умножения: 5x5= 25 и т. п.). Если испытуемый видел цифры умственным взором — значит он «визуальный» тип. а если он хотел произносить соответствующие слова — он «моторный» тип, и т. д. ОБ при таком испытании обнаружили заметно большее внутрипарное сходство, чем РБ. Эти данные служат основанием для вывода о значительной наследственной обусловленности «типа воображения» (Luchsinger, 1944). Здесь самый метод исследования недостаточно объективен, обобщение полученных данных путем установки «типов воображения» недостаточно обосновано и тем более утверждение о наследственности этих типов. И тем не менее в явлениях «внутренней речи» мы имеем дело с реальными фактами, которые надо изучать более объективными методами; и очень вероятно, что и в этом отношении ОБ действительно будут иметь большее внутрипарное сходство, чем РБ. Но здесь важно правильно анализировать эти явления и не спешить говорит о их «наследственности».
Недавно была сделана первая экспериментальная попытка изучения развития речи у близнецов с позиции советской психологии, основывающейся на павловскомпонимании речи как второй сигнальной системы (Лурия и Юдович, 1956). Исследование велось на паре ОБ, Леше и Юре Г., 5 лет, у которых заметно отставало развитие речи. Дети эти родились в срок, их развитие в общем протекало нормально, за исключением речи, и они никаких признаков умственной отсталости или психической дефективности не проявляли. Леша родился 6 фунтов, Юра — 33/4 фунтов; здоровьем Юра был слабее Леши, зубы у него прорезались позднее, чем у его близнеца. Мать считала, что Леша активнее Юры и последний «подчиняется» ему. Однако речевое развитие у близнецов оказалось одинаковым. К 5 годам речь этих детей «состояла из небольшого количества общеупотребительных слов (чаще всего искаженных) и небольшого числа "автономных" слов и звуков»; такая речь выпадала из сложившейся системы русского языка. Это были слова и звуки, вплетенные в непосредственное действие и сопровождающиеся оживленными жестами при общении близнецов друг с другом. Это была «синпрактическая» речь, зависящая только от конкретно-действенной ситуации и очень примитивная. Такая речь сложилась и сохранилась до 5 лет, по-видимому в связи с известной косноязычностью детей и «близнецовой ситуацией» — замкнутостью близнецов в паре, которую они образовывали, — малым общением с окружающим миром. Грамматически полные фразы составляли у близнецов лишь 17.4% их речи, а больше половины высказываний состояло из «аморфных» фраз, содержавших или одно слово или сочетания слов типа «Люля», «Леся», «ага», «нца!». Функциональный анализ речи этих близнецов показал, что «синпрактическая» речь, т. е. сопровождающая конкретные поступки, составляет свыше 90% и до 2.8% речь, выходящая за пределы наглядной ситуации. Чужая речь, выходящая за пределы наглядной ситуации, была непонятна близнецам Г.
Исходя из упомянутых предположений о причинах отсталости развития речи этих близнецов, авторы произвели следующий ряд экспериментов. Чтобы разрушить «близнецовую ситуацию» и вынудить близнецов ближе включиться в детский коллектив (близнецы находились в детском учреждении), а в связи с этим стимулировать развитие нормальной речи, близнецов, очень привязанных друг к другу, отчасти разлучили тем, что поместили в разные группы. Далее, с одним из них, Юрой, дополнительно проводили систематические занятия но обучению речи, развивали у него осознанную речь, привычку пользоваться развернутой фразой и т. д. А Леша служил контролем.
Результаты эксперимента обнаружились быстро. Уже через 3 месяца речь близнецов по своей лексике и грамматике приблизилась к обычной речи их сверстников, хотя и сохранились известные черты прежнего косноязычия. «Аморфные» фразы сократились до 12—19%, а преобладающими стали фразы, понятные вне ситуации (предметные), — 81—88%. Через 10 месяцев «аморфные» фразы вовсе исчезли. Вместе с тем речь Юры, подвергшегося специальному речевому обучению, стала более дифференцированной, с более развернутыми фразами, чем речь Леши, Речь стала для Юры предметом специальной осознанной деятельности, он стал понимать структуру ее, чего Леша не мог. Но особенно существенным оказался тот факт, что одновременно с развитием речи резко изменилась вся структура психической жизни близнецов. Овладев объективной системой языка, дети оказались в состоянии формулировать в речи замысел действия, обнаруживать в связи с этим возможность продуктивной, исходящей из замысла, конструктивной деятельности, развился ряд более сложных интеллектуальных функций, дотоле почти незаметных. Все это очень ясно обнаружилось в играх детей, а также в испытаниях путем вопросов, показа картинок и других приемов.
Эксперимент Лурии и Юдович является несомненно интересным шагом вперед в изучении роли речи в поведении ребенка, возможным только на близнецовом материале.
Дефекты речи, как например косноязычие у вышеописанных близнецов Г., заикание и т. п., у ОБ чаще оказываются конкордатными (Luchsinger, 1940). Это можно толковать как наследственную общность некоторых сторон физиологического механизма этих аномалий, но для того чтобы об этом говорить обоснованно, надо их лучше изучить, пока же они плохо известны.
Рисунки близнецов почти вовсе не изучались. Недавно была сделана попытка психологического толкования таких рисунков (Tisserand-Perrier et Blaizot, 1955). Так, например, если на рисунках пары ОБ, мальчиков 8 лет, один из них, А., изображен тем и другим близнецом с большой головой, то это, по мнению авторов, значит, что оба хотят показать (в соответствии с их высказыванием независимо от этих рисунков), что близнец А. «старше» и «сильнее», т. е. является «ведущим». Так ли надо действительно понимать эти рисунки, остается неясным. На рисунке одной близнячки 22 лет последняя изобразила себя повернутой боком, спиной к своей партнерше. Авторы толкуют этот рисунок как проявление желания рисовавшей быть «независимой» от своей близнячки и т. д. Такие объяснения носят довольно произвольный и не всегда убедительный характер. Нужен метод более строгого и точного анализа материала. Такого метода пока что нет.
Есть указания на существование близнецов-художников (Newman, 1940b). Но их творчество никто не изучал. Не лучше обстоит дело и с изучением музыкальной деятельности близнецов.
Таким образом, и с психологической и с физиологической стороны вторая сигнальная система близнецов, в сущности, почти вовсе не изучена.
Мимика и моторика стали изучаться у близнецов сравнительно недавно и преимущественно с психологических позиций. Впервые систематически этим вопросом занялся Летоваара (Lehtovaara, 1938) на 69 парах детей-близнецов обоих типов. Паре сидящих близнецов показывались поочередно различные картинки смешного или страшного содержания (изувеченное лицо и т. п.), и в это время через отверстие в стенке над демонстрируемой картинкой детей незаметно для них снимали на кинопленку. Кроме того, велся протокол их поведения с записью того, что дети говорили под впечатлением картинок. По фильму изучалось число контактов партнеров, т. е. обращений одного к другому, появление мимического процесса, его тип (различалось 3 типа), особенности и сила. Степень внутрипарного сходства определялась четырьмя баллами. У ОБ контактов оказалось больше, чем у РБ. Тип мимики у ОБ был заметно более одинаков, чем у РБ. Особенно велико было сходство характера мимики у ОБ по сравнению с РБ: 40.8% полной конкордантности у ОБ к 4.3% у РБ. Скорость развития мимического процесса не удалось достаточно точно изучить из-за технических причин (медленность смены кадров). Сходство ОБ оказалось также заметно больше и по силе проявления мимических процессов.
В дальнейшем были поставлены наблюдения еще над 56 парами близнецов обоих типов, в основном в возрасте от 5 до 15 лет (Gedda е Neroni, 1955). Раздражителем были кинофильмы. Из числа нескольких присутствующих на сеансе выборочно снимались отдельные пары фотоаппаратом в трех аспектах (профиль, три четверти и фас), в открытую, т. е. дети могли заметить, что их снимают. Учитывалось положение головы, верхних и нижних конечностей, мимика. Степень внутрипарного сходства оценивалась четырьмя баллами и вычислялся процент оценок по этим баллам к общему числу фотоснимков. Полная конкордантность по мимике в общем обнаружена у 79% ОБ и только у 32.48% РБ, а дискордантность — у 6% ОБ и 29% РБ. Остальное количество падает на промежуточные ступени сходства. Положение головы у ОБ оказалось также более похожим, чем у РБ, и в общем более похожим, чем рук и ног.
На одних взрослых подобные вопросы впервые изучал Сплиндер (Splinder, 1955). Он имел 8 пар ОБ в возрасте от 21 года до 54 лет и 4 пары РБ 20—27 лет внутрипарно одинакового пола, по равному числу пар обоих полов в обеих группах близнецов. Каждая пара росла вместе в относительно одинаковых условиях. Кроме трех пар (из них две РБ), каждая пара имела ту же профессию. Опыты ставились трех типов: 1) изучение мимики (на комическую граммофонную пластинку, на шутки, на веселый фильм и на медицинский фильм — удаление глазного яблока — на «испуг»); 2) изучение «тонкой» моторики (подкраска губ, бритье, вдевание нитки в иголку, писание); 3) изучение «грубой» моторики (бросание и ловля теннисного мяча и т. п.). Испытуемые снимались на кинофильм попарно, иногда и порознь каждый, незаметно для них с помощью особой установки. Зафиксированные в фильме аналогичные реакции каждой пары близнецов, в соответствии с последовательностью раздражений, сравнивались; степень их сходства оценивалась с помощью четырехбалльной системы (конкордантная, дискордантная и две промежуточные ступени) и вычислялся процент оценок к общему числу реакций в пределах всей группы, к которой относится данная пара, т. е. ОБ или РБ. Для мимики получен следующий общий результат в процентах (табл. 30).
В качестве примера резко выраженной «поляризации» взаимоотношений пары ОБ при очень большой взаимной дружбе рассмотрим Наташу и Ноэми (Эму) Ботв., 10 лет (рис. 161). Они родились с очень большим интервалом, почти в 5 дней (имели, по-видимому, разные плаценты и оболочки); с первых месяцев стали проявлять заметное различие характеров: Наташа была более активной по сравнению с Эмой. В раннем детстве они были очень дружны. Как нередко бывает в подобных случаях, их любимым местоимением было «мы», которым они, впрочем, злоупотребляли в некоторых случаях, например, заявив однажды родителям: «Мы сегодня ночью видели во сне. . .», и далее, перебивая друг друга, стали рассказывать содержание сна. Дружба детей не пострадала от того, что более деятельная Наташа стала постепенно инициатором в организации игр, в выполнении различных домашних дел и поручений, представителем близнецов во внешнем мире и т. д., а Эма — ее подсобницей, совершенно пассивной, предоставляющей Наташе активную роль. Это взаимоотношение очень наглядно выступило в одном мелком эпизоде. Эма, во время разговора в лаборатории после опыта, вертела гайку на одном приборе, гайка отлетела и покатилась на пол. Я попросил найти эту гайку. Наташа сразу бросилась искать ее под столом и под соседним шкафом, а Эма, заложив руки за спину спокойно наблюдала за близнячкой. На мой вопрос, почему же ищет Наташа, а не она, отвинтившая гайку. Эма ответила: «Такие дела у нас делает Наташа». При подобном взаимоотношении Эма охотно и покорно подчиняется инициативе Наташи, пользуется ее активностью, а Наташа с удовольствием «командует» и сознает себя незаменимой в их близнецовом коллективе. Степень, которой достигает такого рода «поляризация» у этих близнецов, вредно отзывается на обеих, развивая своеобразную односторонность у каждой из девочек. В таких случаях, как этот, очень ясно выступает значение одного близнеца для другого, как очень действенного и постоянно влияющего фактора среды. С возрастом, с развитием детей такие отношения все больше дифференцируются, пока не достигнут известного предела, также, вероятно, связанного с известной стадией онтогенеза.
Те взаимоотношения, которые так разнообразно слагаются в пределах каждой пары близнецов, особенно любопытно развертываются в близнецовых коллективах с большим числом членов: в тройнях, четвернях и пятернях. Мы рассмотрим два примера: однояйцевую четверню Морлок (Gardner a. Newman, 1943) и однояйцевую пятерню Дионн (Schwesinger, 1940, и др.).
Девочки Морлок (рис. 129) в возрасте 10 лет имели известную разницу в весе и росте. Интересно, что у этих близнецов между величиной роста и веса, с одной стороны, и степенью умственного развития (I.Q.),c другой, была известная корреляция, так же как и у Дионн. Это интересное явление, пока еще недостаточно изученное, могло послужить основой внутренней организации коллектива данной четверни. Заправилой, «лидером» всей группы стала самая крупная близнячка, опережавшая остальных в развитии. Она была представителем интересов четверни в окружающем мире, «депутатом». Самая маленькая в коллективе была «бэби», которую опекали остальные. Две другие взяли на себя роли своего рода затейников: одна была «клоун», задира и шутник, а другая — «артист» в силу большей склонности к рисованию и музыке. Конечно, это не были очень уж прочно фиксированные «роли», но все же они свидетельствовали о внутренней организации коллектива и известном распределении функций между его членами, известной «специализации» их. Морлок пели и танцевали как дружный ансамбль. Интересно, что, в отличие от Дионн, изолированных от внешнего мира, Морлок жили, как прочие дети, в обычном общении со своими сверстниками в школе и вообще с окружающим миром. По умственному развитию они были несколько выше среднего.
У пятерни Дионн (рис. 131), речь о которых подробнее идет в другом месте (глава девятая), внутри их коллектива сложились до известной степени аналогичные отношения: у них также был «лидер» коллектива, была самая «младшая» — «бэби» (Мари), и остальные трое уже с 4 лет проявляли известные индивидуальные особенности. К сожалению, мне неизвестна ни одна работа, в которой бы описывалось развитие индивидуальных особенностей каждой из пяти девочек с раннего возраста (Дионн родились в 1934 г.) в системе их маленького мирка как замкнутого целого.
Одна из попыток проникнуть во внутреннюю структуру коллектива пятерни Дионн принадлежит экологу Элли (Аllее, 1951). В течение 14 месяцев, с 22-го по 36-й месяц жизни детей, наблюдались «контакты» детей друг с другом и велся счет таких соприкосновений. При этом из общего числа различных «контактов» выделялись те из них, когда одни ребенок по своей инициативе адресовался к другому и когда ребенок отвечал на обращение к нему. Отдельные дети из пятерни Дионн заметно отличаются друг от друга в этом отношении, и различия в некоторых случаях носят стойкий и длительный характер. Так, Анетта все время занимала первое место, Цецилия — всегда второе. Остальные имели не столь постоянные места. Между числом контактов и интеллектуальным уровнем (по тестам) корреляции не наблюдалось. Так, самая развитая из группы — Ивонна — по инициативе к контактам занимает последнее место, а «бэби» группы — Мари — среднее место и т. д. Конечно, эти данные Элли лишь очень схематично, односторонне, по аналогии с его опытами с курами, и скудно освещают интересный вопрос структуры такого коллектива и причины этой структуры не раскрывают. Методически изучение этого вопроса должным образом вообще еще не поставлено.
Как одно из проявлений взаимоотношения близнецов надо, по-видимому, рассматривать вопрос об одежде близнецов, хотя он содержит, конечно, и другие элементы. Опрос и наблюдения показали, что пары ОБ чаще, чем пары РБ, желают иметь одинаковую одежду. В этом, очевидно, обнаруживается их чувство солидарности, симпатии друг к другу. Вероятно, что в ряде случаев РБ хотят различием одежды подчеркнуть свою независимость друг от друга, нежелание слыть за близнецов и т. п. (Lehtovaara, 1938; Gedda, 1951, и др.). Однако, как отмечает Гедда, желание иметь одинаковую одежду не всегда совпадает с желанием надевать одежду своего близнеца. Здесь выступает сознание своей индивидуальности, несмотря на симпатию и преданность своему близнецу.
Тот коллектив, который образуют близнецы, особенно если он состоит из большего числа детей, чем двое, иногда приобретает тенденцию замыкаться от окружающего мира, особенно, если близнецы вырабатывают своеобразную речь, понятную только им. Это вредно отражается на развитии близнецов, они отстают от сверстников в смысле психического развития и т. д. (Zazzo, 1952, и др.).
Близнецовый коллектив входит обычно в состав семьи как более крупного коллектива. Влияние отца и матери, братьев и сестер на развитие близнецов, влияние разницы в отношении к близнецам других членов семьи на близнецов и их взаимоотношения — вопросы очень важные для оценки роли социальной среды на формирование психики близнецов. Например, одна из девочек пары ОБ, Нина Ин. о которой речь идет побробнее в другом месте, страдала кривошеей; отец явно больше любил ее нормальную близнячку. Этим, конечно, усиливалось сознание неполноценности у пострадавшей с вытекающими отсюда последствиями. Многообразное и очень важное влияние семьи на развитие психики близнецов еще очень мало исследовано. Примером своеобразного жизненного эксперимента, показывающего важность роли семейной среды, является случай с парой ОБ, описанный Швезингер (Schwesinger, 1952). ОБ Эльвира и Эсфирь росли в разных семьях: первая в семье своей матери, вышедшей вторично замуж и имевшей еще других детей, с которыми Эльвира дружно жила; вторая у суровой приемной матери, в одиночестве и трудных условиях. В материальном отношении обе близнячки жили примерно на одном уровне достаточности. Жизнь Эльвиры сложилась сравнительно удачно, тогда как Эсфирь, замкнутая, неуравновешенная, стала морфинисткой и рано погибла, по-видимому, покончив самоубийством.
Были попытки выяснить социальные отношения близнецов вне семьи — в школьной среде (Lehtovaara, 1938). Это трудная задача, и пока что ее нельзя считать решенной. Выбор детей, с которыми близнецы дружат, совместно играют и т. д., их возраст, пол, число их и проч. — все это учитывалось. Оказалось, что выбор ОБ в отношении товарищей в том же школьном коллективе в общем более похож, чем РБ. Это один из примеров того избирательного отношения к среде, о котором речь была выше.
Социальная ориентация близнецов обнаруживается также в их детских планах относительно будущей профессии. Около 75% пар ОБ имело похожие планы, тогда как только около 50% пар РБ высказало в той или иной мере схожие проекты своего будущего (Lehtovaara, 1938).
Все вышеприведенные данные об отношении близнецов к их социальной среде теснейшим образом связаны с вопросами способов выражения, самообнаружения, общения, т. е. речи, рисунка, мимики, жестикуляции и т. д. К этой группе вопросов мы теперь и переходим.
О развитии речи у близнецов говорится в другом месте (см. главу тринадцатую). С психологических позиций речь близнецов еще очень мало изучена, и физиология речи у близнецов по существу вовсе не изучена. Попытка анализа речи 12 пар близнецов обоих типов по определенной системе (Шейдта), различающей 5 основных классов слов, не обнаружила существенно большего внутрипарного сходства ОБ по сравнению с РБ (Gottschick, 1939). Далее были сделаны попытки установить связь особенностей речи с характеристикой «личности» близнецов и конституциональными типами, моторикой, «личным темпом» и т. д. (Luchsinger, 1940). Все эти попытки малоубедительны с методической и методологической стороны. Например, изучение «внутренней речи» (мыслей и представлений без произнесения слов испытуемым) велось интроспективным методом, т. е. на основании словесного отчета испытуемого после выполнения им определенных заданий (например, «продумать» без произнесения слов 7 пунктов из таблицы умножения: 5x5= 25 и т. п.). Если испытуемый видел цифры умственным взором — значит он «визуальный» тип. а если он хотел произносить соответствующие слова — он «моторный» тип, и т. д. ОБ при таком испытании обнаружили заметно большее внутрипарное сходство, чем РБ. Эти данные служат основанием для вывода о значительной наследственной обусловленности «типа воображения» (Luchsinger, 1944). Здесь самый метод исследования недостаточно объективен, обобщение полученных данных путем установки «типов воображения» недостаточно обосновано и тем более утверждение о наследственности этих типов. И тем не менее в явлениях «внутренней речи» мы имеем дело с реальными фактами, которые надо изучать более объективными методами; и очень вероятно, что и в этом отношении ОБ действительно будут иметь большее внутрипарное сходство, чем РБ. Но здесь важно правильно анализировать эти явления и не спешить говорит о их «наследственности».
Недавно была сделана первая экспериментальная попытка изучения развития речи у близнецов с позиции советской психологии, основывающейся на павловскомпонимании речи как второй сигнальной системы (Лурия и Юдович, 1956). Исследование велось на паре ОБ, Леше и Юре Г., 5 лет, у которых заметно отставало развитие речи. Дети эти родились в срок, их развитие в общем протекало нормально, за исключением речи, и они никаких признаков умственной отсталости или психической дефективности не проявляли. Леша родился 6 фунтов, Юра — 33/4 фунтов; здоровьем Юра был слабее Леши, зубы у него прорезались позднее, чем у его близнеца. Мать считала, что Леша активнее Юры и последний «подчиняется» ему. Однако речевое развитие у близнецов оказалось одинаковым. К 5 годам речь этих детей «состояла из небольшого количества общеупотребительных слов (чаще всего искаженных) и небольшого числа "автономных" слов и звуков»; такая речь выпадала из сложившейся системы русского языка. Это были слова и звуки, вплетенные в непосредственное действие и сопровождающиеся оживленными жестами при общении близнецов друг с другом. Это была «синпрактическая» речь, зависящая только от конкретно-действенной ситуации и очень примитивная. Такая речь сложилась и сохранилась до 5 лет, по-видимому в связи с известной косноязычностью детей и «близнецовой ситуацией» — замкнутостью близнецов в паре, которую они образовывали, — малым общением с окружающим миром. Грамматически полные фразы составляли у близнецов лишь 17.4% их речи, а больше половины высказываний состояло из «аморфных» фраз, содержавших или одно слово или сочетания слов типа «Люля», «Леся», «ага», «нца!». Функциональный анализ речи этих близнецов показал, что «синпрактическая» речь, т. е. сопровождающая конкретные поступки, составляет свыше 90% и до 2.8% речь, выходящая за пределы наглядной ситуации. Чужая речь, выходящая за пределы наглядной ситуации, была непонятна близнецам Г.
Исходя из упомянутых предположений о причинах отсталости развития речи этих близнецов, авторы произвели следующий ряд экспериментов. Чтобы разрушить «близнецовую ситуацию» и вынудить близнецов ближе включиться в детский коллектив (близнецы находились в детском учреждении), а в связи с этим стимулировать развитие нормальной речи, близнецов, очень привязанных друг к другу, отчасти разлучили тем, что поместили в разные группы. Далее, с одним из них, Юрой, дополнительно проводили систематические занятия но обучению речи, развивали у него осознанную речь, привычку пользоваться развернутой фразой и т. д. А Леша служил контролем.
Результаты эксперимента обнаружились быстро. Уже через 3 месяца речь близнецов по своей лексике и грамматике приблизилась к обычной речи их сверстников, хотя и сохранились известные черты прежнего косноязычия. «Аморфные» фразы сократились до 12—19%, а преобладающими стали фразы, понятные вне ситуации (предметные), — 81—88%. Через 10 месяцев «аморфные» фразы вовсе исчезли. Вместе с тем речь Юры, подвергшегося специальному речевому обучению, стала более дифференцированной, с более развернутыми фразами, чем речь Леши, Речь стала для Юры предметом специальной осознанной деятельности, он стал понимать структуру ее, чего Леша не мог. Но особенно существенным оказался тот факт, что одновременно с развитием речи резко изменилась вся структура психической жизни близнецов. Овладев объективной системой языка, дети оказались в состоянии формулировать в речи замысел действия, обнаруживать в связи с этим возможность продуктивной, исходящей из замысла, конструктивной деятельности, развился ряд более сложных интеллектуальных функций, дотоле почти незаметных. Все это очень ясно обнаружилось в играх детей, а также в испытаниях путем вопросов, показа картинок и других приемов.
Эксперимент Лурии и Юдович является несомненно интересным шагом вперед в изучении роли речи в поведении ребенка, возможным только на близнецовом материале.
Дефекты речи, как например косноязычие у вышеописанных близнецов Г., заикание и т. п., у ОБ чаще оказываются конкордатными (Luchsinger, 1940). Это можно толковать как наследственную общность некоторых сторон физиологического механизма этих аномалий, но для того чтобы об этом говорить обоснованно, надо их лучше изучить, пока же они плохо известны.
Рисунки близнецов почти вовсе не изучались. Недавно была сделана попытка психологического толкования таких рисунков (Tisserand-Perrier et Blaizot, 1955). Так, например, если на рисунках пары ОБ, мальчиков 8 лет, один из них, А., изображен тем и другим близнецом с большой головой, то это, по мнению авторов, значит, что оба хотят показать (в соответствии с их высказыванием независимо от этих рисунков), что близнец А. «старше» и «сильнее», т. е. является «ведущим». Так ли надо действительно понимать эти рисунки, остается неясным. На рисунке одной близнячки 22 лет последняя изобразила себя повернутой боком, спиной к своей партнерше. Авторы толкуют этот рисунок как проявление желания рисовавшей быть «независимой» от своей близнячки и т. д. Такие объяснения носят довольно произвольный и не всегда убедительный характер. Нужен метод более строгого и точного анализа материала. Такого метода пока что нет.
Есть указания на существование близнецов-художников (Newman, 1940b). Но их творчество никто не изучал. Не лучше обстоит дело и с изучением музыкальной деятельности близнецов.
Таким образом, и с психологической и с физиологической стороны вторая сигнальная система близнецов, в сущности, почти вовсе не изучена.
Мимика и моторика стали изучаться у близнецов сравнительно недавно и преимущественно с психологических позиций. Впервые систематически этим вопросом занялся Летоваара (Lehtovaara, 1938) на 69 парах детей-близнецов обоих типов. Паре сидящих близнецов показывались поочередно различные картинки смешного или страшного содержания (изувеченное лицо и т. п.), и в это время через отверстие в стенке над демонстрируемой картинкой детей незаметно для них снимали на кинопленку. Кроме того, велся протокол их поведения с записью того, что дети говорили под впечатлением картинок. По фильму изучалось число контактов партнеров, т. е. обращений одного к другому, появление мимического процесса, его тип (различалось 3 типа), особенности и сила. Степень внутрипарного сходства определялась четырьмя баллами. У ОБ контактов оказалось больше, чем у РБ. Тип мимики у ОБ был заметно более одинаков, чем у РБ. Особенно велико было сходство характера мимики у ОБ по сравнению с РБ: 40.8% полной конкордантности у ОБ к 4.3% у РБ. Скорость развития мимического процесса не удалось достаточно точно изучить из-за технических причин (медленность смены кадров). Сходство ОБ оказалось также заметно больше и по силе проявления мимических процессов.
В дальнейшем были поставлены наблюдения еще над 56 парами близнецов обоих типов, в основном в возрасте от 5 до 15 лет (Gedda е Neroni, 1955). Раздражителем были кинофильмы. Из числа нескольких присутствующих на сеансе выборочно снимались отдельные пары фотоаппаратом в трех аспектах (профиль, три четверти и фас), в открытую, т. е. дети могли заметить, что их снимают. Учитывалось положение головы, верхних и нижних конечностей, мимика. Степень внутрипарного сходства оценивалась четырьмя баллами и вычислялся процент оценок по этим баллам к общему числу фотоснимков. Полная конкордантность по мимике в общем обнаружена у 79% ОБ и только у 32.48% РБ, а дискордантность — у 6% ОБ и 29% РБ. Остальное количество падает на промежуточные ступени сходства. Положение головы у ОБ оказалось также более похожим, чем у РБ, и в общем более похожим, чем рук и ног.
На одних взрослых подобные вопросы впервые изучал Сплиндер (Splinder, 1955). Он имел 8 пар ОБ в возрасте от 21 года до 54 лет и 4 пары РБ 20—27 лет внутрипарно одинакового пола, по равному числу пар обоих полов в обеих группах близнецов. Каждая пара росла вместе в относительно одинаковых условиях. Кроме трех пар (из них две РБ), каждая пара имела ту же профессию. Опыты ставились трех типов: 1) изучение мимики (на комическую граммофонную пластинку, на шутки, на веселый фильм и на медицинский фильм — удаление глазного яблока — на «испуг»); 2) изучение «тонкой» моторики (подкраска губ, бритье, вдевание нитки в иголку, писание); 3) изучение «грубой» моторики (бросание и ловля теннисного мяча и т. п.). Испытуемые снимались на кинофильм попарно, иногда и порознь каждый, незаметно для них с помощью особой установки. Зафиксированные в фильме аналогичные реакции каждой пары близнецов, в соответствии с последовательностью раздражений, сравнивались; степень их сходства оценивалась с помощью четырехбалльной системы (конкордантная, дискордантная и две промежуточные ступени) и вычислялся процент оценок к общему числу реакций в пределах всей группы, к которой относится данная пара, т. е. ОБ или РБ. Для мимики получен следующий общий результат в процентах (табл. 30).
 Бросается в глаза внутрипарное сходство мимики ОБ и несходство РБ. Особенно любопытно, что все ОБ реагируют внутрипарно одновременно, все РБ — неодновременно.
При изучении моторики сидящего в кресле испытуемого на том же фильме различали положение корпуса, верхних и нижних конечностей, общее положение. В итоге получены следующие цифры процентов для тех же четырех ступеней сходства (табл. 31).
Бросается в глаза внутрипарное сходство мимики ОБ и несходство РБ. Особенно любопытно, что все ОБ реагируют внутрипарно одновременно, все РБ — неодновременно.
При изучении моторики сидящего в кресле испытуемого на том же фильме различали положение корпуса, верхних и нижних конечностей, общее положение. В итоге получены следующие цифры процентов для тех же четырех ступеней сходства (табл. 31).
 По моторике, сопровождающей мимику и являющейся реакцией на первую группу раздражителей (пластинки и фильмы), внутрипарное сходство ОБ также заметно больше, чем РБ, но в несколько меньшей степени, чем по мимике. То же приблизительно получается и при изучении моторики в других сериях опытов. Однако в серии опытов с преобладанием «произвольных» движений различия между ОБ и РБ почти нет. В проявлении «испуга» (оборонительной реакции на неожиданно с шумом падающий близ испытуемого предмет) часть движений — втягивание шеи и подъем плеч оказалась полностью конкордантной у ОБ и РБ, оборонительное движение рук — почти полностью конкордантным у ОБ и совершенно дискордантным у РБ. Общая поза у ОБ при этом сравнительно очень похожа, а у РБ вовсе непохожа.
Относительно большее внутрипарное сходство ОБ по мимике и моторике, чем РБ, в исследовании Сплиндера по сравнению с данными Гедды объясняется, вероятно, отчасти разницей материала, а также различием в методике, более точной у Сплиндера. Интересна попытка последнего дать физиологическую интерпретацию полученных им данных. Полагая, что иннервация мимических мышц идет от промежуточного мозга экстрапирамидными путями, он считает сходство ОБ в мимике наследственным. Вероятно, в действительности это сложнее, и мимика контролируется корой головного мозга, но, по-видимому, в малой степени, почему у пары ОБ мимика в общем так похожа. В движениях смешанного характера (частью непроизвольных и частью произвольных), иннервация которых идет частично и по пирамидному пути, ОБ внутрипарно меньше похожи, чем в мимике, а в опытах, требующих преимущественно произвольных движений, степень внутрипарного сходства ОБ и РБ оказывается почти одинаковой. Конечно, в такой цепи движений, как например весь процесс бритья, встречаются и непроизвольные движения, и двигательные стереотипы, выработанные известным навыком, и движения, вызванные особенностями данной конкретной обстановки, так сказать, сугубо произвольные в ряде моментов. Все это, конечно, очень усложняет физиологический анализ заснятого процесса бритья и позволяет лишь приблизительно правильно толковать отдельные его звенья.
Наконец, реакция «испуга» в тех сторонах ее проявления, которые всегда похожи у всех ОБ и РБ (втягивание шеи и подъем плеч), объясняется тем, что это движения экстрапирамидной иннервации, свойственные не только людям, но и другим млекопитающим. Иначе говоря, их, по-видимому, надо считать проявлением безусловного оборонительного рефлекса. Дополнительные оборонительные движения рук, туловища и т. д., обусловленные конкретной ситуацией, в известной мере «произвольны», пирамидной иннервации и потому менее внутрипарно похожи у ОБ и в еще меньшей степени у РБ.
При анализе своего материала Сплиндер не пользуется павловскими понятиями. Вероятно, ряд моментов его кинофильма можно было бы лучше интерпретировать в свете учения об условных рефлексах. Экспериментальное изучение мимики, жестикуляции, моторики и т. д. с помощью киносъемки, особенно на детях-близнецах разных возрастов, у нас еще не использованная методика для изучения высшей нервной деятельности.
Недавно сделана попытка изучения моторики ОБ спортсменов, показавшая ряд моментов внутрипарного сходства у них (Grebe, 1955).
По моторике, сопровождающей мимику и являющейся реакцией на первую группу раздражителей (пластинки и фильмы), внутрипарное сходство ОБ также заметно больше, чем РБ, но в несколько меньшей степени, чем по мимике. То же приблизительно получается и при изучении моторики в других сериях опытов. Однако в серии опытов с преобладанием «произвольных» движений различия между ОБ и РБ почти нет. В проявлении «испуга» (оборонительной реакции на неожиданно с шумом падающий близ испытуемого предмет) часть движений — втягивание шеи и подъем плеч оказалась полностью конкордантной у ОБ и РБ, оборонительное движение рук — почти полностью конкордантным у ОБ и совершенно дискордантным у РБ. Общая поза у ОБ при этом сравнительно очень похожа, а у РБ вовсе непохожа.
Относительно большее внутрипарное сходство ОБ по мимике и моторике, чем РБ, в исследовании Сплиндера по сравнению с данными Гедды объясняется, вероятно, отчасти разницей материала, а также различием в методике, более точной у Сплиндера. Интересна попытка последнего дать физиологическую интерпретацию полученных им данных. Полагая, что иннервация мимических мышц идет от промежуточного мозга экстрапирамидными путями, он считает сходство ОБ в мимике наследственным. Вероятно, в действительности это сложнее, и мимика контролируется корой головного мозга, но, по-видимому, в малой степени, почему у пары ОБ мимика в общем так похожа. В движениях смешанного характера (частью непроизвольных и частью произвольных), иннервация которых идет частично и по пирамидному пути, ОБ внутрипарно меньше похожи, чем в мимике, а в опытах, требующих преимущественно произвольных движений, степень внутрипарного сходства ОБ и РБ оказывается почти одинаковой. Конечно, в такой цепи движений, как например весь процесс бритья, встречаются и непроизвольные движения, и двигательные стереотипы, выработанные известным навыком, и движения, вызванные особенностями данной конкретной обстановки, так сказать, сугубо произвольные в ряде моментов. Все это, конечно, очень усложняет физиологический анализ заснятого процесса бритья и позволяет лишь приблизительно правильно толковать отдельные его звенья.
Наконец, реакция «испуга» в тех сторонах ее проявления, которые всегда похожи у всех ОБ и РБ (втягивание шеи и подъем плеч), объясняется тем, что это движения экстрапирамидной иннервации, свойственные не только людям, но и другим млекопитающим. Иначе говоря, их, по-видимому, надо считать проявлением безусловного оборонительного рефлекса. Дополнительные оборонительные движения рук, туловища и т. д., обусловленные конкретной ситуацией, в известной мере «произвольны», пирамидной иннервации и потому менее внутрипарно похожи у ОБ и в еще меньшей степени у РБ.
При анализе своего материала Сплиндер не пользуется павловскими понятиями. Вероятно, ряд моментов его кинофильма можно было бы лучше интерпретировать в свете учения об условных рефлексах. Экспериментальное изучение мимики, жестикуляции, моторики и т. д. с помощью киносъемки, особенно на детях-близнецах разных возрастов, у нас еще не использованная методика для изучения высшей нервной деятельности.
Недавно сделана попытка изучения моторики ОБ спортсменов, показавшая ряд моментов внутрипарного сходства у них (Grebe, 1955).
Глава двенадцатая АНОМАЛИИ И БОЛЕЗНИ У БЛИЗНЕЦОВ
По патологии близнецов собран огромный материал, в ряде случаев еще недостаточно разработанный. Мы ограничимся только немногими вопросами из этой области, выбрав те из них, которые или сравнительно лучше изучены и позволяют делать известные обобщения, или представляют несомненный интерес для нашей проблемы. Начнем с морфологических аномалий. Так, интересны некоторые дефекты лицевого черепа, например щель в верхней челюсти и нёбе, сопровождающаяся также щелью верхней губы, так называемой заячьей губой. Один случай, сравнительно хорошо изученный, демонстративен. Это пара ОБ, Курт и Вальтер, 1909 г. рождения (рис. 162). Щель в губе имелась только у Курта и была зашита, но след ее ясно виден. У Вальтера щели не было. Но на том месте челюсти, где у его близнеца имеется щель, из-за которой у Курта не вырос второй резец, соответствующий второй резец у Вальтера слабо развит — он короче, уже и конической формы. Очевидно, и у Вальтера проявился тот же дефект, но в более ослабленной форме. В роду близнецов эта аномалия раз уже была констатирована: у брата их бабки была волчья пасть и заячья губа. Кроме описанной аномалии, у близнецов установлен еще одинаковый органический недостаток сердца, тоже сильнее выраженный у Курта. Третий дефект обоих — недоразвитие тестикулов, сопровождающееся стерильностью, при в общем нормальном телосложении. У Курта эти железы по размера равны небольшим бобам, у Вальтера крупнее — с косточку сливы (Verschuer, 1954). Возможно, что недоразвитие лицевого черепа как-то связано с недоразвитием гонад у этих близнецов, а также с дефектом сердца. Это остается открытым вопросом (ср. Claussen, 1940). По статистическим данным, заячья губа встречается конкордантно у 33% ОБ и лишь у 12% РБ (Gedda, 1951).
В качестве примера аномалий конечностей рассмотрим интересный случай шестипалости. Это вообще очень редкое уродство. Оно встречается в 0.0025% всех ортопедических аномалий. В литературе известны лишь немногие случаи близнецов с полидактилией, причем в некоторых из них тип близнецов остался невыясненным, что, конечно, снижает ценность этих данных (ср. Gedda, 1951). Мы остановимся в качестве примера только на одной паре ОБ, сравнительно подробно изученной. Это пара девочек на первому году жизни. Шестипалость у них проявилась у обеих, но не вполне конкордантно, как видно из сопоставления внешне наблюдаемой картины (рис. 163)
По статистическим данным, заячья губа встречается конкордантно у 33% ОБ и лишь у 12% РБ (Gedda, 1951).
В качестве примера аномалий конечностей рассмотрим интересный случай шестипалости. Это вообще очень редкое уродство. Оно встречается в 0.0025% всех ортопедических аномалий. В литературе известны лишь немногие случаи близнецов с полидактилией, причем в некоторых из них тип близнецов остался невыясненным, что, конечно, снижает ценность этих данных (ср. Gedda, 1951). Мы остановимся в качестве примера только на одной паре ОБ, сравнительно подробно изученной. Это пара девочек на первому году жизни. Шестипалость у них проявилась у обеих, но не вполне конкордантно, как видно из сопоставления внешне наблюдаемой картины (рис. 163)
 и рентгеновских снимков. У первого близнеца на левой руке имелся дефектный шестой палец — это одна фаланга с ногтем, висящая на стебельке толщиной со спичку и длиной около 0.25 см. (А). Этот палец был лишен подвижности, а также, по-видимому, чувствительности. На правой руке ребенка, на ульнарной стороне мизинца, на верхней трети первой фаланги находится бугорок величиной с булавочную головку — это, вероятно, рудимент шестого пальца (Б). На обеих ногах первого близнеца имеется по шестому пальцу, торчащему вбок (В).
У второго близнеца на левой руке находится вполне развитой шестой палец из трех фаланг, двигающийся одновременно с пятым пальцем (Г). На правой руке, на аналогичном месте, как и у первого близнеца, имелся продолговатый выступ с утолщением на конце, толщиной с булавку и длиной около 0.25 см. Первая фаланга мизинца, на которой находился этот вырост, была толще нормальной фаланги. Левая нога этого ребенка несла на себе шестой палец, как соответственная нога первого ребенка. Правая нога второго близнеца шестого пальца не имела, но пятый был крупнее нормы и торчал в сторону. Если для наглядности сопоставить эти данные в виде таблицы, то получим следующую картину (табл. 32).
и рентгеновских снимков. У первого близнеца на левой руке имелся дефектный шестой палец — это одна фаланга с ногтем, висящая на стебельке толщиной со спичку и длиной около 0.25 см. (А). Этот палец был лишен подвижности, а также, по-видимому, чувствительности. На правой руке ребенка, на ульнарной стороне мизинца, на верхней трети первой фаланги находится бугорок величиной с булавочную головку — это, вероятно, рудимент шестого пальца (Б). На обеих ногах первого близнеца имеется по шестому пальцу, торчащему вбок (В).
У второго близнеца на левой руке находится вполне развитой шестой палец из трех фаланг, двигающийся одновременно с пятым пальцем (Г). На правой руке, на аналогичном месте, как и у первого близнеца, имелся продолговатый выступ с утолщением на конце, толщиной с булавку и длиной около 0.25 см. Первая фаланга мизинца, на которой находился этот вырост, была толще нормальной фаланги. Левая нога этого ребенка несла на себе шестой палец, как соответственная нога первого ребенка. Правая нога второго близнеца шестого пальца не имела, но пятый был крупнее нормы и торчал в сторону. Если для наглядности сопоставить эти данные в виде таблицы, то получим следующую картину (табл. 32).
 В семье больше ни у кого шестипалости не установлено. Но обнаружились небольшие аномалии мизинцев (малый размер и ограниченная подвижность) у бабушки близнецов по материнской линии, у матери их, у одной из сестер матери, (тетки близнецов) и у сестры этих детей (Lehmann u. Wittler, 1935). Трудно допустить, чтобы шестипалость у этих близнецов была случайным совпадением при той редкости этой аномалии, которая известна. Вероятно, у них имелось какое-то особое генотипическое предрасположение к шестипалости. Картина в этом отношении не ясна. Эта гипотетическая предрасположенность неодинаково проявлялась на разных конечностях в смысле степени выраженности признака: от утолщения пятого пальца и бугорка на мизинце до развитого шестого пальца из трех фаланг. Эта изменчивость проявления признака, вероятно, зависит от условий утробного развития близнецов, пока что тоже еще невыясненных. Мы видим, что этот любопытный случай шестипалости у ОБ вызывает целую группу вопросов, которые предстоит решать будущим исследователям.
В семье больше ни у кого шестипалости не установлено. Но обнаружились небольшие аномалии мизинцев (малый размер и ограниченная подвижность) у бабушки близнецов по материнской линии, у матери их, у одной из сестер матери, (тетки близнецов) и у сестры этих детей (Lehmann u. Wittler, 1935). Трудно допустить, чтобы шестипалость у этих близнецов была случайным совпадением при той редкости этой аномалии, которая известна. Вероятно, у них имелось какое-то особое генотипическое предрасположение к шестипалости. Картина в этом отношении не ясна. Эта гипотетическая предрасположенность неодинаково проявлялась на разных конечностях в смысле степени выраженности признака: от утолщения пятого пальца и бугорка на мизинце до развитого шестого пальца из трех фаланг. Эта изменчивость проявления признака, вероятно, зависит от условий утробного развития близнецов, пока что тоже еще невыясненных. Мы видим, что этот любопытный случай шестипалости у ОБ вызывает целую группу вопросов, которые предстоит решать будущим исследователям.
 Здесь надо отметить, что шестипалость обнаружена еще у некоторых пар близнецов. Так, у пары соединенных близнецов (рис. 58) на всех конечностях было по 6 пальцев (рис. 164). Еще у одной пары ОБ было по лишнему пальцу рядом с мизинцем на каждой руке, а на ногах имелось нормальное количество пальцев (Ruhl, 1938). Далее, из пары близнецов женского пола (тип близнецов не указан), лишь у одной из близнячек была шестипалость, притом очень определенного типа: на левой руке рядом с мизинцем был лишний палец из трех фаланг. Удивительно, что эта аномалия прослежена в роду этих близнецов в 6 поколениях и в одной семье этого рода, где оба супруга имели эту аномалию, все 8 детей ее тоже имели, и все только на левой руке (Barrer, 1947). Здесь, по-видимому, шестипалость носит несколько иной наследственный характер, чем в описанном выше случае Лемана. Наконец, найден был редчайший случай шестипалости в виде удвоения большого пальца и притом только у одного из ОБ, которые оба имели раздвоенный позвоночник — spina bifida (Newman a. Quisenberry, 1944).
Здесь надо отметить, что шестипалость обнаружена еще у некоторых пар близнецов. Так, у пары соединенных близнецов (рис. 58) на всех конечностях было по 6 пальцев (рис. 164). Еще у одной пары ОБ было по лишнему пальцу рядом с мизинцем на каждой руке, а на ногах имелось нормальное количество пальцев (Ruhl, 1938). Далее, из пары близнецов женского пола (тип близнецов не указан), лишь у одной из близнячек была шестипалость, притом очень определенного типа: на левой руке рядом с мизинцем был лишний палец из трех фаланг. Удивительно, что эта аномалия прослежена в роду этих близнецов в 6 поколениях и в одной семье этого рода, где оба супруга имели эту аномалию, все 8 детей ее тоже имели, и все только на левой руке (Barrer, 1947). Здесь, по-видимому, шестипалость носит несколько иной наследственный характер, чем в описанном выше случае Лемана. Наконец, найден был редчайший случай шестипалости в виде удвоения большого пальца и притом только у одного из ОБ, которые оба имели раздвоенный позвоночник — spina bifida (Newman a. Quisenberry, 1944).
 Из других аномалий рук мы кратко рассмотрим еще один случай: синдактилии — соединения пальцев (рис. 165), вместе с контрактурой четвертого и пятого пальца у пары ОБ (Bauer, 1927). Сходство здесь настолько велико, что говорить о случайном совпадении не приходится. Любопытно, что во время операции разъединения пальцев была сделана пересадка кожи с одного близнеца на другого, вполне удачная, чего обычно при пересадке кожи с одного субъекта на другого не бывает. Этот факт дополняет опыты с пересадкой кожи у близнецов-коров (см. глава десятая «Кожа»).
Из многочисленных аномалий нижних конечностей рассмотрим еще только косолапость. В серии без отбора установлены (Verschuer, 1952) следующие цифровые отношения (табл. 33).
Из других аномалий рук мы кратко рассмотрим еще один случай: синдактилии — соединения пальцев (рис. 165), вместе с контрактурой четвертого и пятого пальца у пары ОБ (Bauer, 1927). Сходство здесь настолько велико, что говорить о случайном совпадении не приходится. Любопытно, что во время операции разъединения пальцев была сделана пересадка кожи с одного близнеца на другого, вполне удачная, чего обычно при пересадке кожи с одного субъекта на другого не бывает. Этот факт дополняет опыты с пересадкой кожи у близнецов-коров (см. глава десятая «Кожа»).
Из многочисленных аномалий нижних конечностей рассмотрим еще только косолапость. В серии без отбора установлены (Verschuer, 1952) следующие цифровые отношения (табл. 33).
 Эти цифры говорят о заметно большем проценте конкордантных пар среди ОБ по сравнению с РБ, у которых относительное число косолапых приблизительно так же велико, как у страдающих этим дефектом обычных братьев-сестер близнецов. Процент конкордантных пар ОБ не велик (22.9), что говорит о сравнительно небольшой генотипической тенденции к развитию косолапости. на проявление болезни влияют как внутренние факторы, например пол (так как косолапых мужчин вдвое больше, чем женщин), так и различные внешние факторы среды, географические и иные, способствующие или препятствующие развитию косолапости. В общем же роль среды как и генотипа в этом вопросе еще недостаточно изучена.
Эти цифры говорят о заметно большем проценте конкордантных пар среди ОБ по сравнению с РБ, у которых относительное число косолапых приблизительно так же велико, как у страдающих этим дефектом обычных братьев-сестер близнецов. Процент конкордантных пар ОБ не велик (22.9), что говорит о сравнительно небольшой генотипической тенденции к развитию косолапости. на проявление болезни влияют как внутренние факторы, например пол (так как косолапых мужчин вдвое больше, чем женщин), так и различные внешние факторы среды, географические и иные, способствующие или препятствующие развитию косолапости. В общем же роль среды как и генотипа в этом вопросе еще недостаточно изучена.
Болезни кожи и ее производных
Многочисленные дефекты и болезни кожи были изучены у близнецов. Обзор этих работ дает Гедда (Gedda, 1951). Мы можем здесь в качестве примеров привести лишь немногие факты. Так, любопытен случай большой конкордантности по заболеванию угрями (акне) у пары ОБ. Как видно из рис. 166, пятна этой сыпи на лице расположены не на гомологичных местах у обоих близнецов; однако на левом мизинце у той и другой на одном и том же месте имеется дерматофиброма (Melsom, 1945). Псориаз (чешуйчатый лишай), этиология которого не выяснена, описан по крайней мере у 20 пар ОБ, из них 12 пар конкордантны, 8 — дискордантны. У одной пары 8-летних однояйцевых мальчиков конкордантный псориаз оказался на довольно необычном месте — на спине. Кроме того, псориаз имелся у них еще на голове. Болезнь у одного обнаружилась на год раньше, чем у другого. У отца их псориаз появился в возрасте 18 лет (Gedda, 1951). Роль наследственности при псориазе еще недостаточно изучена (ср. Vogel, 1956).
Альбинизм, встречающийся среди населения США примерно один случай на тысячу человек, описан был у пары ОБ (рис. 167). Такой случай можно ожидать в среднем одни на 3 миллиона человек. Как типичные альбиносы, эти близнецы оба имели слабое зрение. Все их родственники имели нормальную пигментацию (Rife et al., 1946).
Псориаз (чешуйчатый лишай), этиология которого не выяснена, описан по крайней мере у 20 пар ОБ, из них 12 пар конкордантны, 8 — дискордантны. У одной пары 8-летних однояйцевых мальчиков конкордантный псориаз оказался на довольно необычном месте — на спине. Кроме того, псориаз имелся у них еще на голове. Болезнь у одного обнаружилась на год раньше, чем у другого. У отца их псориаз появился в возрасте 18 лет (Gedda, 1951). Роль наследственности при псориазе еще недостаточно изучена (ср. Vogel, 1956).
Альбинизм, встречающийся среди населения США примерно один случай на тысячу человек, описан был у пары ОБ (рис. 167). Такой случай можно ожидать в среднем одни на 3 миллиона человек. Как типичные альбиносы, эти близнецы оба имели слабое зрение. Все их родственники имели нормальную пигментацию (Rife et al., 1946).

Аномалии желез внутренней секреции
Расстройства эндокринного аппарата, как известно, оказывают в некоторых случаях очень глубокое влияние на весь процесс развития человека, на всю его жизнь. Из уже довольно большого материала по эндокринным заболеваниям у близнецов (см. Gedda, 1951) мы можем здесь лишь кратко остановится на немногих, наиболее показательных случаях. Так, например, пара ОБ, Анна и Мария, развивались очень различно (рис. 168). Анна рано стала отставать от Марин; уже с 3-летнего возраста разница стала очень заметна, особенно же резко она выступала в период полового созревания (Luth, 1937). Причина предполагалась в расстройстве функции гипофиза у Анны, возникшем еще в период утробного развития; близнецы родились на восьмом месяце беременности. Другой случай резкого различия в развитии пары ОБ 14 лет возник на почве базедовой болезни (расстройство функции щитовидной железы) у одной из близнячек, отставшей из-за этого от другой, нормальной. Чем вызвано заболевание только одной из пары, остается неясным (Borchardt, 1931). В литературе описан ряд случаев базедовой болезни у близнецов, как ОБ, так и РБ. По-видимому, среди ОБ чаще встречаются конкордантные, чем дискордантные пары, как вышеописанная, тогда как среди РБ преобладают дискордантные пары. Пары женского пола приблизительно в 3 раза чаще болеют базедовой болезнью, чем мужчины (Lehmann, 1938).
Эндемичный зоб (гипертрофия щитовидной железы) на исключительно большом материале — 520 пар близнецов — исследовал Эйгстер (Eugster, 1936) у швейцарского населения. Как известно, в некоторых местностях Швейцарии зоб частое явление. Зависимость возникновения зоба от места обитания и ряда конкретных условий среды уже раньше была установлена. Поэтому Эйгстер очень тщательно изучал условия жизни обследуемых близнецов у них дома, входя в разные подробности: местность, этаж квартиры, род пищи, занятие и т. д. В связи с этим он установил пять степеней сходства-различия условий среды. Оказалось, что у близнецов обоих типов, при одинаковых условиях среды для каждой пары, все же наблюдаются внутрипарная разница в развитии щитовидной железы, что в ряде случаев зависит от определенных различий в обстоятельствах жизни в прошлом. Отношение числа конкордантных пар к дискордантным в процентах для 148 пар с одинаковой средой получилось следующее (табл. 34).
Другой случай резкого различия в развитии пары ОБ 14 лет возник на почве базедовой болезни (расстройство функции щитовидной железы) у одной из близнячек, отставшей из-за этого от другой, нормальной. Чем вызвано заболевание только одной из пары, остается неясным (Borchardt, 1931). В литературе описан ряд случаев базедовой болезни у близнецов, как ОБ, так и РБ. По-видимому, среди ОБ чаще встречаются конкордантные, чем дискордантные пары, как вышеописанная, тогда как среди РБ преобладают дискордантные пары. Пары женского пола приблизительно в 3 раза чаще болеют базедовой болезнью, чем мужчины (Lehmann, 1938).
Эндемичный зоб (гипертрофия щитовидной железы) на исключительно большом материале — 520 пар близнецов — исследовал Эйгстер (Eugster, 1936) у швейцарского населения. Как известно, в некоторых местностях Швейцарии зоб частое явление. Зависимость возникновения зоба от места обитания и ряда конкретных условий среды уже раньше была установлена. Поэтому Эйгстер очень тщательно изучал условия жизни обследуемых близнецов у них дома, входя в разные подробности: местность, этаж квартиры, род пищи, занятие и т. д. В связи с этим он установил пять степеней сходства-различия условий среды. Оказалось, что у близнецов обоих типов, при одинаковых условиях среды для каждой пары, все же наблюдаются внутрипарная разница в развитии щитовидной железы, что в ряде случаев зависит от определенных различий в обстоятельствах жизни в прошлом. Отношение числа конкордантных пар к дискордантным в процентах для 148 пар с одинаковой средой получилось следующее (табл. 34).
 Степень конкордантности ОБ и РБо почти одинакова. Это говорит в пользу предположения, что в основном эндемичный зоб зависит от условий среды. В пользу этого говорит и то, что ОБ с внутрипарно разной средой имеют приблизительно в 7 раз больше различий, чем ОБ с одинаковой средой. Особенно велико внутрипарное различие, если один из близнецов живет в местности, где зоб распространен, т. е., по-видимому, подвергнут определенным химическим воздействиям через питьевую воду и т. д. Для патогенеза зоба и возникновения внутрипарной разницы большое значение имеет состояние щитовидной железы при рождении ребенка, зависящее от негенетических факторов утробной жизни. Нет никакого наследственного предрасположения к зобу. В генетическом смысле эта болезнь — модификация. Генотип в ней может сказаться лишь в возможностях реакции организма на становление зоба, что выражается в ходе процесса развития заболевания, локализации и патологоанатомической формы зоба. Поэтому у РБ внутрипарно несколько чаще встречаются качественные различия заболевания.
Нам остается еще сказать несколько слов об эндокринной функции гонад. Недоразвитие мужских половых желез (тестикулов) сочетается иногда с крипторхизмом — расположением этих желез в брюшной полости вместо мошонки. Такой конкордантный случай у пары ОБ сопровождался отставанием в развитии. Более отстававший из двух по росту и весу на 13-м году жизни подвергся лечению путем введения гонадотропного гормона, а второй близнец служил контролем. Через 8 месяцев лечения эффект был заметный: ранее отстававший в развитии теперь обогнал своего близнеца (контрольного) на 5 см по росту и оказался на 3.6 кг тяжелее, у него появились признаки половой зрелости, голос стал низкий и зычный и т. д. Контрольный близнец остался таким же отсталым, с детским голосом (рис. 169). Возмужавший близнец перешел в следующий класс, а его близнец (не лечившийся) остался в том же классе на второй год (Greene, 1942).
Степень конкордантности ОБ и РБо почти одинакова. Это говорит в пользу предположения, что в основном эндемичный зоб зависит от условий среды. В пользу этого говорит и то, что ОБ с внутрипарно разной средой имеют приблизительно в 7 раз больше различий, чем ОБ с одинаковой средой. Особенно велико внутрипарное различие, если один из близнецов живет в местности, где зоб распространен, т. е., по-видимому, подвергнут определенным химическим воздействиям через питьевую воду и т. д. Для патогенеза зоба и возникновения внутрипарной разницы большое значение имеет состояние щитовидной железы при рождении ребенка, зависящее от негенетических факторов утробной жизни. Нет никакого наследственного предрасположения к зобу. В генетическом смысле эта болезнь — модификация. Генотип в ней может сказаться лишь в возможностях реакции организма на становление зоба, что выражается в ходе процесса развития заболевания, локализации и патологоанатомической формы зоба. Поэтому у РБ внутрипарно несколько чаще встречаются качественные различия заболевания.
Нам остается еще сказать несколько слов об эндокринной функции гонад. Недоразвитие мужских половых желез (тестикулов) сочетается иногда с крипторхизмом — расположением этих желез в брюшной полости вместо мошонки. Такой конкордантный случай у пары ОБ сопровождался отставанием в развитии. Более отстававший из двух по росту и весу на 13-м году жизни подвергся лечению путем введения гонадотропного гормона, а второй близнец служил контролем. Через 8 месяцев лечения эффект был заметный: ранее отстававший в развитии теперь обогнал своего близнеца (контрольного) на 5 см по росту и оказался на 3.6 кг тяжелее, у него появились признаки половой зрелости, голос стал низкий и зычный и т. д. Контрольный близнец остался таким же отсталым, с детским голосом (рис. 169). Возмужавший близнец перешел в следующий класс, а его близнец (не лечившийся) остался в том же классе на второй год (Greene, 1942).
 Явления псевдогермафродитизма описаны у ОБ, как конкордатные (Rhodes, 1943; Collier, 1948), так и дискордантные (Guldberg, 1938) в очень редких случаях (всего 10 пар).
Гомосексуализм многократно изучался у близнецов (Gedda, 1951). В этих явлениях трудно различить биологическую основу от социальных влияний. Конкордантность в разной степени найдена у большинства OБ (Kallman, 1952а).
Явления псевдогермафродитизма описаны у ОБ, как конкордатные (Rhodes, 1943; Collier, 1948), так и дискордантные (Guldberg, 1938) в очень редких случаях (всего 10 пар).
Гомосексуализм многократно изучался у близнецов (Gedda, 1951). В этих явлениях трудно различить биологическую основу от социальных влияний. Конкордантность в разной степени найдена у большинства OБ (Kallman, 1952а).
Заболевания нервной системы
Многие нервные и психические заболевания изучались у близнецов (ср.: Gedda, 1951; Slater, 1953, и др.). Здесь невозможно рассматривать весь этот огромный материал. Мы остановимся только на двух болезнях, в которых наследственность, судя по ряду исследований, играет известную роль. Это шизофрения (раннее слабоумие) и эпилепсия. По шизофрении у близнецов опубликован ряд работ (Luxenburger, 1930; Rosanoff a. Tuman-Kane, 1934; Essen-Moller, 1941; Kallmann, 1946, 1950, 1952b; Slater, 1953, и др.), охватывающих очень большой материал. Мы остановимся кратко лишь на работах двух исследователей — Коллмана и Слэтера. Коллман собрал 953 пары близнецов, где хотя бы один из партнеров страдал шизофренией. Это была безотборная серия, однако близнецы брались только в возрасте свыше 15 лет. Каждый близнец был полностью обследован, а кроме того, различные родственники близнецов — родители и братья-сестры (БС) — в общем свыше 6000 человек. Коллман сравнил относительное число больных не только среди обоих типов близнецов (ОБ и РБ), но также среди упомянутых групп родственников, т. е. работал расширенным близнецовым методом «методом близнецовой семьи» («twin family method»), который он применял также при изучении других болезней, в частности туберкулеза (см. далее «Туберкулез»). Результаты сведены в схему (рис. 170). Мы видим, что процент больных детей у больных родителей заметно больше, чем у здоровых родителей, и особенно велик, когда оба родителя страдают шизофренией. Далее видно, что среди сводных братьев-сестер близнецов, лиц, не имеющих с ними кровного родства, процент шизофреников очень мал, он увеличивается у полу-БС (братьев-сестер, имеющих одного общего родителя), он еще больше у настоящих братьев-сестер близнецов и одинаков с цифрой процента у РБ. Однако у ОБ процент больных партнеров больного близнеца достигает мaксимума, особенно же он велик у близнецов, воспитанных вместе, т. е. имеющих схожую среду и влияющих друг на друга (например, больной близнец влияет на заболевающего и т. п.).
Эти данные приводят Коллмана к выводу, что наследственность играет несомненную роль в стойкости организма в отношении шизофрении, чем, конечно, вопрос не решается. Это лишь приближение к постановке изучения физиологического механизма возникновения и развития заболевания, а в связи с этим и его лечения. Углубленное изучение отдельных пар на протяжении возможно длительного периода времени может дать ценный материал для этой цели, в частности сравнение пары ОБ в том случае, когда один из близнецов болеет, а другой здоров. Например, одна из близнячек подобной пары, болевшая шизофренией, поправилась в связи с увеличением веса, превзойдя свою здоровую партнершу на 22 фунта. Коллман думает, что подобные наблюдения открывают пути для профилактики и лечения шизофрении. В ряде случаев отмечается удивительное сходство возникновения и развития заболевания у пары ОБ, что толкуется как наследственное сходство по нестойкости нервной системы (Gardner a. Stephens, 1949).
Слэтер собрал меньший материал (41 пару ОБ и соответственно меньшее число РБ и БС, чем Коллман). Среди ОБ было 76% конкордантных пар, при 14% среди РБ и только 5% среди БС. Возможно, как считает сам автор, процент БС занижен из-за менее полного изучения последних. Течение болезни и исход ее у конкордантных пар ОБ были далеко не всегда одинаковы, что говорит о значительной роли факторов среды. В рассмотрение казуистики, конкретизирующей этот вывод, мы пускаться здесь не можем.
Эпилепсия многократно изучалась у близнецов (см. Gedda, 1951). Крупнейшей по объему работой последнего времени является исследование Леннокса с сотрудниками (Lennox et al., 1945; Lennox, 1951, 1956), собравшего 122 пары припадочных близнецов, помимо большого материала по эпилептикам-одиночкам и их родным (всего свыше 24 000 человек). Данные по близнецам (Lennox, 1951) сведены в табл. 35; словами «мимолетные припадки» здесь обозначены разные слабые проявления эпилепсии.
Мы видим, что процент больных детей у больных родителей заметно больше, чем у здоровых родителей, и особенно велик, когда оба родителя страдают шизофренией. Далее видно, что среди сводных братьев-сестер близнецов, лиц, не имеющих с ними кровного родства, процент шизофреников очень мал, он увеличивается у полу-БС (братьев-сестер, имеющих одного общего родителя), он еще больше у настоящих братьев-сестер близнецов и одинаков с цифрой процента у РБ. Однако у ОБ процент больных партнеров больного близнеца достигает мaксимума, особенно же он велик у близнецов, воспитанных вместе, т. е. имеющих схожую среду и влияющих друг на друга (например, больной близнец влияет на заболевающего и т. п.).
Эти данные приводят Коллмана к выводу, что наследственность играет несомненную роль в стойкости организма в отношении шизофрении, чем, конечно, вопрос не решается. Это лишь приближение к постановке изучения физиологического механизма возникновения и развития заболевания, а в связи с этим и его лечения. Углубленное изучение отдельных пар на протяжении возможно длительного периода времени может дать ценный материал для этой цели, в частности сравнение пары ОБ в том случае, когда один из близнецов болеет, а другой здоров. Например, одна из близнячек подобной пары, болевшая шизофренией, поправилась в связи с увеличением веса, превзойдя свою здоровую партнершу на 22 фунта. Коллман думает, что подобные наблюдения открывают пути для профилактики и лечения шизофрении. В ряде случаев отмечается удивительное сходство возникновения и развития заболевания у пары ОБ, что толкуется как наследственное сходство по нестойкости нервной системы (Gardner a. Stephens, 1949).
Слэтер собрал меньший материал (41 пару ОБ и соответственно меньшее число РБ и БС, чем Коллман). Среди ОБ было 76% конкордантных пар, при 14% среди РБ и только 5% среди БС. Возможно, как считает сам автор, процент БС занижен из-за менее полного изучения последних. Течение болезни и исход ее у конкордантных пар ОБ были далеко не всегда одинаковы, что говорит о значительной роли факторов среды. В рассмотрение казуистики, конкретизирующей этот вывод, мы пускаться здесь не можем.
Эпилепсия многократно изучалась у близнецов (см. Gedda, 1951). Крупнейшей по объему работой последнего времени является исследование Леннокса с сотрудниками (Lennox et al., 1945; Lennox, 1951, 1956), собравшего 122 пары припадочных близнецов, помимо большого материала по эпилептикам-одиночкам и их родным (всего свыше 24 000 человек). Данные по близнецам (Lennox, 1951) сведены в табл. 35; словами «мимолетные припадки» здесь обозначены разные слабые проявления эпилепсии.
 Мы должны прежде всего обратить внимание на число пар ОБ и РБ. Последних не только относительно меньше, чем в популяции, но даже и абсолютно меньше, чем ОБ. Это затрудняет обычное в таких случаях сравнение и говорит за известный отбор однояйцевых пар.
Возраст близнецов был от 6 месяцев до 42 лет. Материал рассматривается без учета возрастных особенностей проявления эпилепсии.
В таблице различаются пары с повреждением мозга (ушиб и т. п.) и без такого повреждения. Внутрипарная конкордантность у пар ОБ без повреждения значительно больше (84%), чем у пар с повреждением (17%, — см. последнюю графу табл. 35), что Леннокс толкует как проявление генотипической предпосылки эпилепсии.
Особый интерес представляют днскордантные случаи ОБ; их 7 из 45 пар ОБ, что составляет 16%. В каждой из этих пар страдает припадками только один из близнецов. Интересно, что, например, у одного не болевшего 12 лет близнеца из числа этих 7 пар во время сна удалось получить электроэнцефалограмму с признаками, свойственными эпилептикам. Тщательное изучение аномалий электроволн мозга может показать предрасположение к эпилептическому припадку задолго до его появления. Вообще же электроэнцефалограммы ОБ-эпилептиков так же внутрипарно похожи, как электроэнцефалограммы здоровых ОБ (см. глава десятая «Нервная система»), конечно с некоторыми исключениями.
Среди РБ было 9% (5 из 53) конкордантных пар. Эта цифра более чем в 2 раза превышает процент случаев эпилепсии (3.4) среди 8006 братьев-сестер изучавшейся серии эпилептиков. Почему у эпилептика его близнец из другого яйца болеет более чем в 2 раза чаще, чем просто брат или сестра эпилептика, пока непонятно. Может быть это зависит от того сходства близнецов вообще, о котором говорит Гедда (глава десятая «Нервная система»).
Форма проявления эпилепсии, как это видно из данных табл. 35, также заметно более конкордантна внутрипарно у ОБ, чем у РБ; например, мы видим, что хроническими припадками страдает из числа ОБ без повреждения мозга 47%, а среди РБ — только 2.5%, и т. д. Клиническая картина, электроэнцефалограммы свидетельствуют о том же. Это говорит о несомненной роли генотипа в заболевании эпилепсией. Леннокс не пытается ближе определить эту роль. Очевидно, его материал не дает к тому основания. Новейшее исследование по генетике эпилепсии (Kimball a. Hersh, 1955) показывает, что этот вопрос требует, конечно, дальнейшего изучения.
Внутрипарное сравнение умственных способностей 87 пар эпилептиков с помощью тестов показало, что кроме лиц, имевших травму мозга (ушиб и т. д.), интеллект эпилептиков, с известными колебаниями, не ниже средней нормы (Lennox а. Collins, 1945).
Мы должны прежде всего обратить внимание на число пар ОБ и РБ. Последних не только относительно меньше, чем в популяции, но даже и абсолютно меньше, чем ОБ. Это затрудняет обычное в таких случаях сравнение и говорит за известный отбор однояйцевых пар.
Возраст близнецов был от 6 месяцев до 42 лет. Материал рассматривается без учета возрастных особенностей проявления эпилепсии.
В таблице различаются пары с повреждением мозга (ушиб и т. п.) и без такого повреждения. Внутрипарная конкордантность у пар ОБ без повреждения значительно больше (84%), чем у пар с повреждением (17%, — см. последнюю графу табл. 35), что Леннокс толкует как проявление генотипической предпосылки эпилепсии.
Особый интерес представляют днскордантные случаи ОБ; их 7 из 45 пар ОБ, что составляет 16%. В каждой из этих пар страдает припадками только один из близнецов. Интересно, что, например, у одного не болевшего 12 лет близнеца из числа этих 7 пар во время сна удалось получить электроэнцефалограмму с признаками, свойственными эпилептикам. Тщательное изучение аномалий электроволн мозга может показать предрасположение к эпилептическому припадку задолго до его появления. Вообще же электроэнцефалограммы ОБ-эпилептиков так же внутрипарно похожи, как электроэнцефалограммы здоровых ОБ (см. глава десятая «Нервная система»), конечно с некоторыми исключениями.
Среди РБ было 9% (5 из 53) конкордантных пар. Эта цифра более чем в 2 раза превышает процент случаев эпилепсии (3.4) среди 8006 братьев-сестер изучавшейся серии эпилептиков. Почему у эпилептика его близнец из другого яйца болеет более чем в 2 раза чаще, чем просто брат или сестра эпилептика, пока непонятно. Может быть это зависит от того сходства близнецов вообще, о котором говорит Гедда (глава десятая «Нервная система»).
Форма проявления эпилепсии, как это видно из данных табл. 35, также заметно более конкордантна внутрипарно у ОБ, чем у РБ; например, мы видим, что хроническими припадками страдает из числа ОБ без повреждения мозга 47%, а среди РБ — только 2.5%, и т. д. Клиническая картина, электроэнцефалограммы свидетельствуют о том же. Это говорит о несомненной роли генотипа в заболевании эпилепсией. Леннокс не пытается ближе определить эту роль. Очевидно, его материал не дает к тому основания. Новейшее исследование по генетике эпилепсии (Kimball a. Hersh, 1955) показывает, что этот вопрос требует, конечно, дальнейшего изучения.
Внутрипарное сравнение умственных способностей 87 пар эпилептиков с помощью тестов показало, что кроме лиц, имевших травму мозга (ушиб и т. д.), интеллект эпилептиков, с известными колебаниями, не ниже средней нормы (Lennox а. Collins, 1945).
Туберкулез
Вопрос о роли факторов наследственности и среды в этиологии этой болезни, ее развития и исходе уже давно интересовал науку. Привлечение близнецового материала могло помочь в решении этой важной проблемы. Из работ по изучению туберкулеза у близнецов основополагающими являются две монографии Диля и Фершюра (Diehl u. Verschuer, 1933, 1936), в которых опубликованы результаты многолетнего исследования этих авторов, а также монография Мичриха (Mitschrich, 1955), в которой обобщаются результаты обследования тех же больных через 20 и более лет после первого обследования. Основное исследование сделано на 205 парах близнецов обоих типов, у которых изучался преимущественно легочный туберкулез, а также костный и другие формы. Больные были главным образом взрослые. Диль и Фершюр на основании этих данных делают вывод, что преобладание конкордантности у ОБ надо объяснять существованием наследственной «тенденции к туберкулезу». Большее внутрипарное сходство ОБ, чем РБ, становится наглядным при конкретном изучении возникновения, локализации и течения болезни у отдельных пар близнецов. Авторы отмечают ряд случаев, когда внутрипарное сходство заболевания ОБ наблюдалось несмотря на различие условий жизни. В качестве примера последнего приводится, в частности, такой случай. Две сестры ОБ 26 лет в течение 9 лет жили в разных местах и имели разные профессии. Одна была продавщицей в Берлине, другая — швеей в сельской местности в Восточной Пруссии. Обе почти одновременно заболели тяжелой формой туберкулеза: у обеих обнаружена каверна в левом легком под ключицей. Обе погибли, причем берлинская близнячка, которую стали лечить на 3 месяца раньше, чем ее деревенскую сестру, пережила последнюю на 5 месяцев. Толкование этого случая как сходства заболевания даже при разных условиях жизни можно принять лишь с оговоркой следующего рода: хотя обе близнячки жили в разных географических пунктах Германии, имели разные профессии и проч., но в основном условия их жизни все же были, вероятно, похожи. Сходство заключалось в том, что они жили в очень неблагоприятных для них условиях, от чего обе и погибли. Работа Диля и Фершюра вызвала ряд возражений, прежде всего методического характера в смысле оценки одинаковой среды, отнесения отдельных пар в рубрики конкордантных и т. д. (Lange, 1935; Rossle, 1940; Roeder, 1950, и др.). Мы не можем входить здесь в рассмотрение этой полемики. Остановимся лишь на фактах Босика (1934), данные которого, полученные на московских детях, не могут, как считает сам Босик, вполне подтвердить выводы Диля и Фершюра. Босик изучил две формы заболевания туберкулезом, чаще всего встречающиеся у детей, — туберкулезную интоксикацию и туберкулезный бронхаденит. Из 130 пар обследованных ОБ 63.7% было здоровых, 21.5% конкордантных по туберкулезной интоксикации и 10.8% конкордантных по бронхадениту, всего 96.0% конкордантных и только 4.0% дискордантных пар ОБ, т. е. таких, где один был здоров, а другой болен, или один страдал одной формой туберкулеза, а другой иной. Среди РБо дискордантных пар было 11.8%, а среди РБр — 8.3%, т. е. дискордантность явно большая, чем среди ОБ. Босик считает, однако, что превалирующую роль в этиологии этих форм туберкулеза играют факторы среды, а наследственные факторы, по-видимому, имеют незначительное влияние. Свое расхождение с выводами Диля и Фершюра Босик объясняет, с одной стороны, неодинаковостью материала обоих исследований, а с другой стороны, допускает возможность изменения соотносительной роли наследственности и среды в зависимости от той конкретной среды, в которой живут исследуемые. Эти соображения, конечно, нуждаются в дальнейшей проверке на новом материале и в специальном исследовании влияния возраста на туберкулез и различные формы его у близнецов. Данные Диля и Фершюра нашли известное подтверждение в позднейших исследованиях (Verschuer, 1955), как это видно из табл. 36.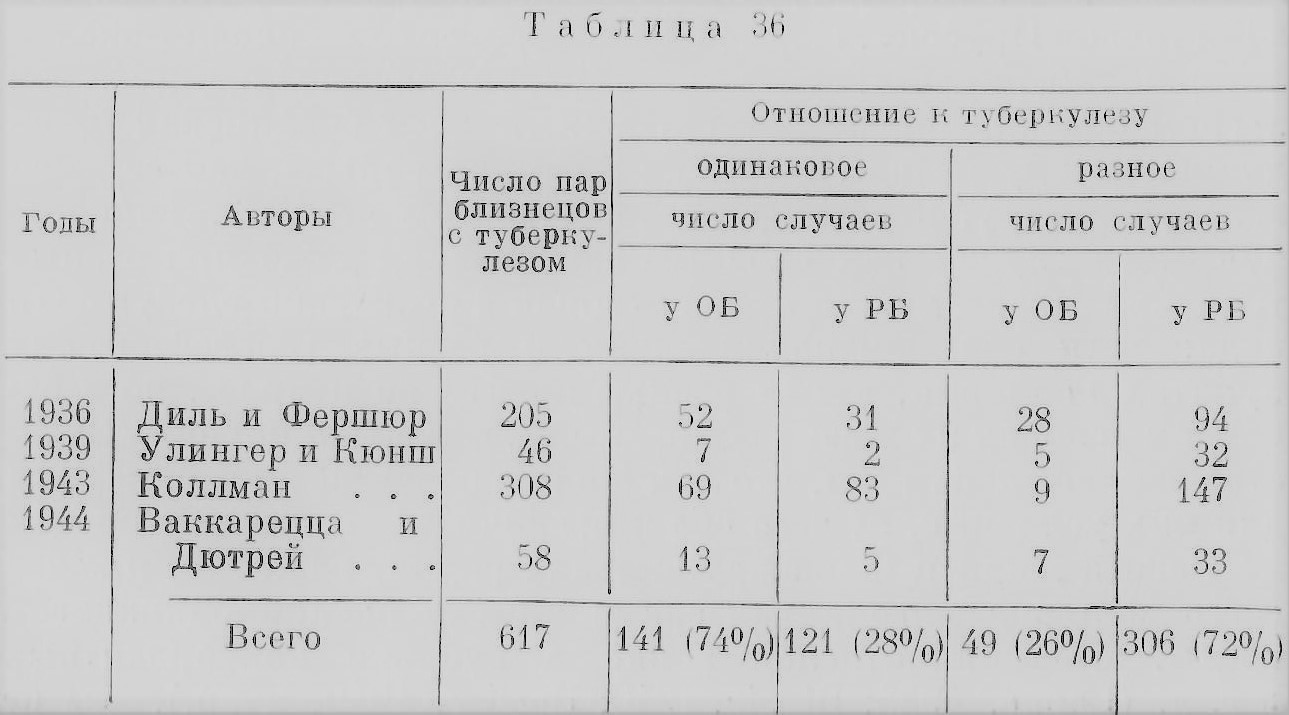 Из этих цифр Фершюр делает вывод: «Очевидно, что наследственное предрасположение оказывает существенное влияние на отношение человека к туберкулезу».
Было опубликовано продолжение работы Диля и Фершюра: 118 пар близнецов из прежде изученных этими авторами (в 1935 г.) было повторно обследовано через 20 и более лет после их первоначального исследования (Mitschrich, 1955; Verschuer, 1955). У 102 пар из них можно было констатировать либо развитие специфического туберкулезного процесса, либо прекращение первоначального процесса. Результаты прежнего (раньше) и нового (теперь) обследования приведены в табл. 37.
Из этих цифр Фершюр делает вывод: «Очевидно, что наследственное предрасположение оказывает существенное влияние на отношение человека к туберкулезу».
Было опубликовано продолжение работы Диля и Фершюра: 118 пар близнецов из прежде изученных этими авторами (в 1935 г.) было повторно обследовано через 20 и более лет после их первоначального исследования (Mitschrich, 1955; Verschuer, 1955). У 102 пар из них можно было констатировать либо развитие специфического туберкулезного процесса, либо прекращение первоначального процесса. Результаты прежнего (раньше) и нового (теперь) обследования приведены в табл. 37.
 Статистически достоверной разницы между отношением к туберкулезу раньше и теперь у тех же пар близнецов установить нельзя. Кажется даже, что конкордантность у ОБ и дискордантность у РБ несколько увеличилась со временем.
Интересна судьба близнецов, выясняющаяся при дальнейшем наблюдении, как это видно из табл. 38.
Статистически достоверной разницы между отношением к туберкулезу раньше и теперь у тех же пар близнецов установить нельзя. Кажется даже, что конкордантность у ОБ и дискордантность у РБ несколько увеличилась со временем.
Интересна судьба близнецов, выясняющаяся при дальнейшем наблюдении, как это видно из табл. 38.

В дискордантных парах ОБ, т. е. таких, в которых болел только один из близнецов, большинство выздоровело, а в дискордантных парах РБ много смертей. Среди конкордантных пар с ранней формой туберкулеза много выздоровевших как РБ, так и ОБ. При поздней форме четверо из ранее легко болевших ОБ умерло вслед за умершим партнером, чего у РБ не наблюдалось. Теперь нам надо остановиться на работе Коллмана и Рейснера (Kallmann a. Reisner, 1943), самой большой по объему охваченного материала среди работ по туберкулезу у близнецов. Она сделана на населении штата и города Нью-Йорка. Брались без отбора все случаи близнецов, где хоть один из пары болел туберкулезом, притом прошедшие через одну из туберкулезных больниц или клиник и получивших там соответствующий диагноз. Кроме самих близнецов, в исследование привлекались члены их семей различных степеней родства. Это делалось по следующим методическим соображениям: чтобы выяснить значение генотипа для заболевания и развития туберкулеза, интересно было сравнить ОБ не только с РБ, но также с лицами, в разной мере далеко стоящими друг от друга в смысле сходства но генотипу. Таким образом, обычный близнецовый метод соединяется с изучением семьи близнецов («метод близнецовой семьи»), У пары родителей различали шесть категорий потомства, которые можно сравнивать: ОБ, РБ одинакового пола, РБ разного пола, полные БС, т е. братья-сестры, полу-БС (имеющие одного общего родителя) и, наконец, сводные братья-сестры (не имеющие кровного родства между собой). Кроме того, для сравнения привлекались по мере надобности родители и супруги близнецов. В этом исследовании участвовало несколько туберкулезных учреждений, работа продолжалась около 5 лет. Собранный материал состоял из 308 семей близнецов, имевших 616 человек близнецов; из них было 78 пар ОБ и 230 пар РБ, из числа которых 118 пар одного пола и 112 разного. Количественное соотношение обоих типов близнецов в этом материале приблизительно соответствует их отношению в популяции. Кроме того, 930 полных БС, 74 полу-БС, 688 родителей, 226 супругов близнецов, т. е. их жен или мужей, всего 2534 человека было исследовано в отношении туберкулеза и контакта с больными им. Основной возраст исследуемых был 15—29 лет. Возрастные колебания устранялись путем соответствующего расчета по методу Вейнберга (Weinberg, 1930).
 Сопоставление числа заболевших в каждой из вышеуказанных групп в семьях больных близнецов с общим числом лиц в каждой из этих групп показало долю больных в процентах. Эти цифры выражены в виде диаграммы (рис. 171, А). Заболеваемость обоих партнеров ОБ почти в 3 раза больше заболеваемости партнеров РБ одинакового пола (87.3 к 30.2). Заболеваемость РБ стоит на одном уровне с БС и выше, чем заболеваемость других родственников. Заболеваемость супругов значительно выше заболеваемости в популяции, что объясняется большим контактом с больным, однако заметно ниже заболеваемости родственников, что, по-видимому, зависит от генотипа.
Для выяснения роли контакта с больными особо изучались семьи с больными родителями и семьи со здоровыми родителями.Если один из родителей или оба болеют туберкулезом, то процент больных БС и РБ возрастает почти вдвое по сравнению с семьями, где родители здоровы (рис. 171, Б). ОБ почти в одинаковой мере, притом в большей, чем РБ, могут стать больными независимо от здоровья родителей в отношении туберкулеза. Удивительно, что заболеваемость ОБ в семьях, где родители здоровы, заметно превышает заболеваемость РБ в семьях с больными родителями. По-видимому, это можно объяснить только большим внутрипарным генотипическим сходством ОБ по сравнению с РБ. Эти данные дополняются сведениями относительно близнецов, о которых известно, что они вообще подвергались возможности заразиться туберкулезом, и теми, о которых это неизвестно. Заболеваемость среди РБ, подвергавшихся такой возможности, больше, чем среди РБ, ей, видимо, не подвергавшихся (рис. 172); однако среди ОБ заболеваемость оказывается в обоих случаях почти одинаковой.
Интересно, что заболеваемость «небелых» людей в материале американских авторов примерно такаяже, как и белых, однако «фатальный» исход приблизительно вдвое больше у первых по сравнению с последними. Авторы усматривают в этом проявление «расового фактора», забывая о социально-экономическом, который здесь, вероятно, играет решающую роль. Вообще роль социально-экономических факторов почти вовсе не освещена в данном исследовании.
Большое внутрипарное сходство ОБ выступает также при сравнении развития и исхода заболевания туберкулезом. Вариации резистентности к туберкулезу, определяемые по внутрипарной разнице в степени и ходе развития заболевания приведены в табл. 39, где клиническое состояние близнецов поделено на 4 подгруппы (I —IV).
Сопоставление числа заболевших в каждой из вышеуказанных групп в семьях больных близнецов с общим числом лиц в каждой из этих групп показало долю больных в процентах. Эти цифры выражены в виде диаграммы (рис. 171, А). Заболеваемость обоих партнеров ОБ почти в 3 раза больше заболеваемости партнеров РБ одинакового пола (87.3 к 30.2). Заболеваемость РБ стоит на одном уровне с БС и выше, чем заболеваемость других родственников. Заболеваемость супругов значительно выше заболеваемости в популяции, что объясняется большим контактом с больным, однако заметно ниже заболеваемости родственников, что, по-видимому, зависит от генотипа.
Для выяснения роли контакта с больными особо изучались семьи с больными родителями и семьи со здоровыми родителями.Если один из родителей или оба болеют туберкулезом, то процент больных БС и РБ возрастает почти вдвое по сравнению с семьями, где родители здоровы (рис. 171, Б). ОБ почти в одинаковой мере, притом в большей, чем РБ, могут стать больными независимо от здоровья родителей в отношении туберкулеза. Удивительно, что заболеваемость ОБ в семьях, где родители здоровы, заметно превышает заболеваемость РБ в семьях с больными родителями. По-видимому, это можно объяснить только большим внутрипарным генотипическим сходством ОБ по сравнению с РБ. Эти данные дополняются сведениями относительно близнецов, о которых известно, что они вообще подвергались возможности заразиться туберкулезом, и теми, о которых это неизвестно. Заболеваемость среди РБ, подвергавшихся такой возможности, больше, чем среди РБ, ей, видимо, не подвергавшихся (рис. 172); однако среди ОБ заболеваемость оказывается в обоих случаях почти одинаковой.
Интересно, что заболеваемость «небелых» людей в материале американских авторов примерно такаяже, как и белых, однако «фатальный» исход приблизительно вдвое больше у первых по сравнению с последними. Авторы усматривают в этом проявление «расового фактора», забывая о социально-экономическом, который здесь, вероятно, играет решающую роль. Вообще роль социально-экономических факторов почти вовсе не освещена в данном исследовании.
Большое внутрипарное сходство ОБ выступает также при сравнении развития и исхода заболевания туберкулезом. Вариации резистентности к туберкулезу, определяемые по внутрипарной разнице в степени и ходе развития заболевания приведены в табл. 39, где клиническое состояние близнецов поделено на 4 подгруппы (I —IV).
 Из табл. 39 видно, что характер течения болезни, в котором обнаруживается резистентность к туберкулезу, очень различается у обоих типов близнецов. Если у ОБ несходство резистентности к сходству относится как 2 : 16 (9 и 69 пар), то у РБ соответственное отношение будет 2 : 1 (147 и 83 пары). Иначе говоря, похожее течение болезни у ОБ встречается приблизительно в 8 раз чаще, чем непохожее, и только в 4 раза чаще у разнояйцевых партнеров. Таким образом, различие между ОБ и РБ по резистентности в процессе хода болезни еще больше, чем по заболеваемости. Отчего это зависит, остается невыясненным.
Этот вопрос и некоторые, с ним связанные, еще недостаточно разработаны и требуют дополнительных исследований (клинического материала, антропометрического и т. д.), как это признают и сами авторы. До сих пор, однако, насколько мне известно, такие материалы не опубликованы.
Недостаток клинического материала в работе Коллмана отчасти восполняется недавним исследованием Плананского и Аллена (Planansky a. Allen, 1953). Эти авторы изучили 43 пары ОБ и 30 пар РБ, все конкордантные в смысле заболевания, т. е. во всех этих парах оба близнеца болели хроническим легочным туберкулезом. Эти больные находились под наблюдением в течение 10 лет. Кроме того, изучались еще 40 пар ОБ с одним больным партнером. Авторов интересовало начало, течение и исход болезни, а также различные, влияющие на нее факторы, в частности госпитализация. Влияние последней на одного или обоих партнеров показывают данные табл. 40.
Из табл. 39 видно, что характер течения болезни, в котором обнаруживается резистентность к туберкулезу, очень различается у обоих типов близнецов. Если у ОБ несходство резистентности к сходству относится как 2 : 16 (9 и 69 пар), то у РБ соответственное отношение будет 2 : 1 (147 и 83 пары). Иначе говоря, похожее течение болезни у ОБ встречается приблизительно в 8 раз чаще, чем непохожее, и только в 4 раза чаще у разнояйцевых партнеров. Таким образом, различие между ОБ и РБ по резистентности в процессе хода болезни еще больше, чем по заболеваемости. Отчего это зависит, остается невыясненным.
Этот вопрос и некоторые, с ним связанные, еще недостаточно разработаны и требуют дополнительных исследований (клинического материала, антропометрического и т. д.), как это признают и сами авторы. До сих пор, однако, насколько мне известно, такие материалы не опубликованы.
Недостаток клинического материала в работе Коллмана отчасти восполняется недавним исследованием Плананского и Аллена (Planansky a. Allen, 1953). Эти авторы изучили 43 пары ОБ и 30 пар РБ, все конкордантные в смысле заболевания, т. е. во всех этих парах оба близнеца болели хроническим легочным туберкулезом. Эти больные находились под наблюдением в течение 10 лет. Кроме того, изучались еще 40 пар ОБ с одним больным партнером. Авторов интересовало начало, течение и исход болезни, а также различные, влияющие на нее факторы, в частности госпитализация. Влияние последней на одного или обоих партнеров показывают данные табл. 40.
 У ОБ исход болезни в большой мере зависит от госпитализации. Вообще легочный туберкулез протекает у ОБ внутрипарно более похоже, чем у РБ. Так, например, из 43 пар ОБ у 23 пар начальное состояние внутрипарно было очень похоже, тогда как из 30 пар РБ это наблюдалось только у 7 пар. Вместе с тем резистентность разных пар ОБ к туберкулезу оказалась неодинакова. Авторы приходят к выводу, что резистентность определяется мультифакториальным генетическим механизмом, который находится в значительной зависимости от неспецифической наследственности (других факторов генотипа) и негенетических факторов (среды, других болезней и т. п.).
Этим, конечно, нельзя считать вопрос о роли генотипа при туберкулезе решенным. Длительные наблюдения над близнецами, клинические и неклинические, могут дать для этого ценный материал.
В недавно опубликованной работе, сделанной на английском населении (Simonds, 1957), показано, что процент случаев заболевания обоих близнецов одной пары сравнительно мал, а в смысле сходства заболевания ОБ приблизительно на 10% превосходят РБ. Интерпретация этих данных недостаточна.
Применение близнецового метода в туберкулезной клинике, помимо чисто теоретического значения, может дать и лечебный эффект. Почин в этом направлении принадлежит советским ученым, начавшим изучать результат лечения кварцевой лампой на близнецах (Босик с сотрудниками, 1934, 1936).
В первой серии опытов использовано 9 пар ОБ, во второй — 13 пар. Выбраны такие дети, у которых клиническая картина заболевания была внутрипарно сходна. В каждой из пар один близнец служил для экспериментального облучения, другой в качестве контроля. Исследование показало, что курс лечения кварцевой лампой имел положительный эффект непосредственно после облучения (была заметна прибавка в весе облученных по сравнению с контрольными). Однако действие облучения было непродолжительным. Улучшения состояния бронхиальных желез под влиянием кварцевой лампы не наблюдалось. Терапевтических преимуществ от увеличения или уменьшения дозировки облучения установить не удалось. Однако меньшая доза способствовала большей прибавке веса непосредственно после облучения.
Другой опыт применения метода взаимоконтроля близнецов был опубликован недавно Монди (Mondy, 1953). У пары девочек ОБ 5 лет имелся туберкулезный процесс, одинаково локализованный в правом легком. У одной из близнячек заболевание осложнилось еще плевритом. Ее одну и стали лечить стрептомицином, как более тяжело болевшую. Для обеих якобы стрептомицина не хватало. Улучшение наступило раньше и заметнее у этой девочки, что, по мнению автора, служит доказательством эффективности лечения легочного туберкулеза стрептомицином. Разумеется, что этим методом можно и следует глубже изучать как этот лечебный прием, так и различные другие.
У ОБ исход болезни в большой мере зависит от госпитализации. Вообще легочный туберкулез протекает у ОБ внутрипарно более похоже, чем у РБ. Так, например, из 43 пар ОБ у 23 пар начальное состояние внутрипарно было очень похоже, тогда как из 30 пар РБ это наблюдалось только у 7 пар. Вместе с тем резистентность разных пар ОБ к туберкулезу оказалась неодинакова. Авторы приходят к выводу, что резистентность определяется мультифакториальным генетическим механизмом, который находится в значительной зависимости от неспецифической наследственности (других факторов генотипа) и негенетических факторов (среды, других болезней и т. п.).
Этим, конечно, нельзя считать вопрос о роли генотипа при туберкулезе решенным. Длительные наблюдения над близнецами, клинические и неклинические, могут дать для этого ценный материал.
В недавно опубликованной работе, сделанной на английском населении (Simonds, 1957), показано, что процент случаев заболевания обоих близнецов одной пары сравнительно мал, а в смысле сходства заболевания ОБ приблизительно на 10% превосходят РБ. Интерпретация этих данных недостаточна.
Применение близнецового метода в туберкулезной клинике, помимо чисто теоретического значения, может дать и лечебный эффект. Почин в этом направлении принадлежит советским ученым, начавшим изучать результат лечения кварцевой лампой на близнецах (Босик с сотрудниками, 1934, 1936).
В первой серии опытов использовано 9 пар ОБ, во второй — 13 пар. Выбраны такие дети, у которых клиническая картина заболевания была внутрипарно сходна. В каждой из пар один близнец служил для экспериментального облучения, другой в качестве контроля. Исследование показало, что курс лечения кварцевой лампой имел положительный эффект непосредственно после облучения (была заметна прибавка в весе облученных по сравнению с контрольными). Однако действие облучения было непродолжительным. Улучшения состояния бронхиальных желез под влиянием кварцевой лампы не наблюдалось. Терапевтических преимуществ от увеличения или уменьшения дозировки облучения установить не удалось. Однако меньшая доза способствовала большей прибавке веса непосредственно после облучения.
Другой опыт применения метода взаимоконтроля близнецов был опубликован недавно Монди (Mondy, 1953). У пары девочек ОБ 5 лет имелся туберкулезный процесс, одинаково локализованный в правом легком. У одной из близнячек заболевание осложнилось еще плевритом. Ее одну и стали лечить стрептомицином, как более тяжело болевшую. Для обеих якобы стрептомицина не хватало. Улучшение наступило раньше и заметнее у этой девочки, что, по мнению автора, служит доказательством эффективности лечения легочного туберкулеза стрептомицином. Разумеется, что этим методом можно и следует глубже изучать как этот лечебный прием, так и различные другие.
Другие инфекции
Не входя в детальное рассмотрение других заразных болезней мы лишь кратко остановимся на некоторых цифровых данных, показывающих конкордантность—дискордантность различных заболеваний у обоих типов близнецов. Первая из приводимых таблиц (табл. 41) заимствована из статьи Глатцеля (Glatzel, 1931), в которой использован материал о заболеваемости среди немцев. Дискордантными считались те случаи, когда болен только один из близнецов или болел в другое время, конкордантными — когда болели оба одновременно или почти одновременно, причем клиническая картина заболевания была похожа. Условия среды для каждой пары «одинаковые».
Первая из приводимых таблиц (табл. 41) заимствована из статьи Глатцеля (Glatzel, 1931), в которой использован материал о заболеваемости среди немцев. Дискордантными считались те случаи, когда болен только один из близнецов или болел в другое время, конкордантными — когда болели оба одновременно или почти одновременно, причем клиническая картина заболевания была похожа. Условия среды для каждой пары «одинаковые».
 Данные табл. 42 взяты из работы Босика (1934), сделанной на материале русских детей.
Картина по обеим таблицам получается довольно пестрая. Большее внутрипарное сходство ОБ нигде резко не выступает. Босик отмечает лишь значительно большую дискордантность РБ, чем ОБ, при воспалении легких. Но по данным Глатцеля такой разницы нет. Большая конкордантность в таких случаях, как при кори, например, может скорее зависеть от заразительности болезни, чем от генотипических свойств заболевшего. Несколько большее внутрипарное сходство течения болезни у ОБ по сравнению с РБ дает право говорить о небольшом значении генотипа в реакции организма больного на инфекцию (Босик, 1934). Фершюр (Verschuer, 1937), собрав еще больший материал по этим болезням, на основании сопоставления коэффициентов конкордантности для разных болезней у обоих типов близнецов, утверждал, что генотип в известной доле участвует в этиологии некоторых из этих болезней, например кори, инфлуенцы и других. Однако этим вопрос еще нельзя считать решенным; он требует дальнейшего изучения (Gedda, 1951). Это относится и к разным другим заразным болезням, на которых мы здесь не будем останавливаться. Новые данные, говорят, по-видимому, о малой роли генотипа в реакции на инфекцию.
Данные табл. 42 взяты из работы Босика (1934), сделанной на материале русских детей.
Картина по обеим таблицам получается довольно пестрая. Большее внутрипарное сходство ОБ нигде резко не выступает. Босик отмечает лишь значительно большую дискордантность РБ, чем ОБ, при воспалении легких. Но по данным Глатцеля такой разницы нет. Большая конкордантность в таких случаях, как при кори, например, может скорее зависеть от заразительности болезни, чем от генотипических свойств заболевшего. Несколько большее внутрипарное сходство течения болезни у ОБ по сравнению с РБ дает право говорить о небольшом значении генотипа в реакции организма больного на инфекцию (Босик, 1934). Фершюр (Verschuer, 1937), собрав еще больший материал по этим болезням, на основании сопоставления коэффициентов конкордантности для разных болезней у обоих типов близнецов, утверждал, что генотип в известной доле участвует в этиологии некоторых из этих болезней, например кори, инфлуенцы и других. Однако этим вопрос еще нельзя считать решенным; он требует дальнейшего изучения (Gedda, 1951). Это относится и к разным другим заразным болезням, на которых мы здесь не будем останавливаться. Новые данные, говорят, по-видимому, о малой роли генотипа в реакции на инфекцию.
Рак
Не касаясь здесь данных относительно различных опухолей у близнецов (обзор дает Гедда), мы остановимся только на раке. Вопрос о роли наследственности при этом заболевании не раз ставился в литературе, и были многократные попытки выяснить его путем изучения рака у близнецов (Реуron et Kobozieff, 1937; Habs, 1938, 1939; Macklin, 1940; Verschuer u. Kober, 1940; Мартынова, 1945; Schinz et al., 1948; Mey, 1950; Charache, 1953, и др.). He раз высказывалось мнение, что существует известное наследственное предрасположение к раку, при этом для обоснования его иногда приводился отобранный материал или отдельные «интересные» случаи. Фершюр и Кобер (Verschuer u. Kober, 1940) собрали одну из первых серий по раку без отбора материала и констатировали относительно очень малую внутрипарную конкордантность, как видно из табл. 43, в которой сопоставлены данные этой серии с результатами исследований нескольких других ученых (Verschuer, 1952).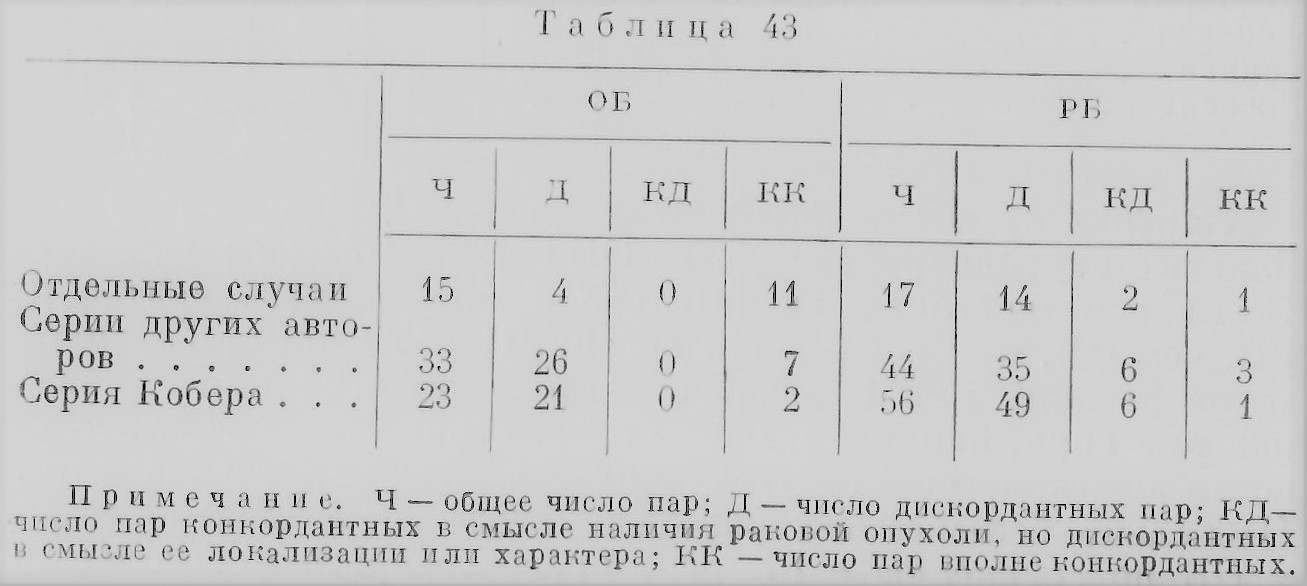 В материале Кобера бросается в глаза большое число дискордантных пар ОБ, лишь сравнительно немного меньшее, чем число дискордантных пар РБ. В материале других авторов, особенно среди отдельных случаев, дискордантных пар ОБ относительно меньше.
Через 12 лет Кобер вновь исследовал своих раковых близнецов. Из 21 дискордантной пары ОБ 2 стали конкордантными: у обеих пар оказался рак желудка. Из 49 дискордантных пар РБ тоже 2 пары стали конкордантными, но одна оказалась не вполне конкордантной: у одной близнячки был рак груди, у другой — прямой кишки. В общем же преобладание дискордантных пар ОБ сохранилось в этой серии и через 12 лет. Две особенности группы конкордантных однояйцевых пар, отличающие их от разнояйцевых, надо отметить: число вполне конкордантных пар все же в 4—5 раз больше среди ОБ, чем среди РБ, — это во-первых, а во-вторых, среди ОБ нет вовсе пар частично конкордантных, какие встречаются среди РБ, и этих случаев в 6 раз больше среди близнецов этого типа, чем вполне конкордатных. Это говорит о том, что у ОБ место и характер опухоли внутрипарно одинаковы в том случае, когда рак есть у обоих близнецов. У РБ преобладает обратная картина — различие локализации и характера опухоли. Данные эти в общем подтверждаются наблюдениями на 7 парах близнецов, обследованных через 25 лет Фершюром (Verschuer, 1954).
Полученные Фершюром и Кобером данные можно толковать следующим образом. Преобладание дискордантных пар среди ОБ свидетельствует о том, что генотипические факторы играют в возникновении и развитии рака незначительную роль, поэтому говорить о наследственности рака не приходится. Однако известную роль генотип при заболевании раком все же играет, что видно из разницы конкордантности среди ОБ и РБ. Роль генотипа обнаруживается во внутрипарном сходстве ОБ в смысле локализации и хода процесса заболевания, до известной степени аналогично тому, что мы видели при туберкулезе.
Фершюр не знал работы Мартыновой (1945), собравшей еще больший материал без отбора по раку у близнецов. Опубликована была только часть данных — о 31 паре ОБ и 95 парах РБ (всего 126 пар). Из пар ОБ только 4 пары оказались конкордантными, т. е. болели оба близнеца,что составляло 12.9%. Среди РБ было 13.7 % конкордантных пар (13 пар), т. е. в сущности то же относительное количество, как и среди ОБ. На основании этого Мартынова считает, что если в этиологии рака наследственность и играет какую-то роль, то не основную.
Что касается сходства локализации раковой опухоли, то из 4 пар ОБ, болевших раком желудочно-кишечного тракта, полное внутрипарное сходство установлено у 3 пар, т. е. у 75%. Из 22 пар РБ внутрипарное сходство найдено только у одной пары (рак желудка у обоих близнецов), т. е. в 4.5%. Это огромная разница между обоими типами близнецов. Если она будет установлена на большем материале с полной достоверностью, то толковать ее надо в том смысле, что локализация рака в большой мере обусловлена генотипом.
Внутрипарное сходство ОБ обычно не ограничивается только местом образования опухоли. Оно распространяется и на возраст появления опухоли и на гистологическую структуру ее. Пара ОБ не бывает частично похожа, только по локализации, например, или только по структуре опухоли, тогда как у РБ внутрипарное сходство ограничивается только наличием опухоли у обоих близнецов, с различием в ряде других моментов. Поразительное сходство заболевания раком ОБ отмечали и другие исследователи (ср. Gedda, 1951). Таким образом, если в возникновении рака генотип играет, по-видимому, незначительную роль, то на течение заболевания, локализацию и строение опухоли и т. д. генотип оказывает весьма существенное влияние.
В материале Кобера бросается в глаза большое число дискордантных пар ОБ, лишь сравнительно немного меньшее, чем число дискордантных пар РБ. В материале других авторов, особенно среди отдельных случаев, дискордантных пар ОБ относительно меньше.
Через 12 лет Кобер вновь исследовал своих раковых близнецов. Из 21 дискордантной пары ОБ 2 стали конкордантными: у обеих пар оказался рак желудка. Из 49 дискордантных пар РБ тоже 2 пары стали конкордантными, но одна оказалась не вполне конкордантной: у одной близнячки был рак груди, у другой — прямой кишки. В общем же преобладание дискордантных пар ОБ сохранилось в этой серии и через 12 лет. Две особенности группы конкордантных однояйцевых пар, отличающие их от разнояйцевых, надо отметить: число вполне конкордантных пар все же в 4—5 раз больше среди ОБ, чем среди РБ, — это во-первых, а во-вторых, среди ОБ нет вовсе пар частично конкордантных, какие встречаются среди РБ, и этих случаев в 6 раз больше среди близнецов этого типа, чем вполне конкордатных. Это говорит о том, что у ОБ место и характер опухоли внутрипарно одинаковы в том случае, когда рак есть у обоих близнецов. У РБ преобладает обратная картина — различие локализации и характера опухоли. Данные эти в общем подтверждаются наблюдениями на 7 парах близнецов, обследованных через 25 лет Фершюром (Verschuer, 1954).
Полученные Фершюром и Кобером данные можно толковать следующим образом. Преобладание дискордантных пар среди ОБ свидетельствует о том, что генотипические факторы играют в возникновении и развитии рака незначительную роль, поэтому говорить о наследственности рака не приходится. Однако известную роль генотип при заболевании раком все же играет, что видно из разницы конкордантности среди ОБ и РБ. Роль генотипа обнаруживается во внутрипарном сходстве ОБ в смысле локализации и хода процесса заболевания, до известной степени аналогично тому, что мы видели при туберкулезе.
Фершюр не знал работы Мартыновой (1945), собравшей еще больший материал без отбора по раку у близнецов. Опубликована была только часть данных — о 31 паре ОБ и 95 парах РБ (всего 126 пар). Из пар ОБ только 4 пары оказались конкордантными, т. е. болели оба близнеца,что составляло 12.9%. Среди РБ было 13.7 % конкордантных пар (13 пар), т. е. в сущности то же относительное количество, как и среди ОБ. На основании этого Мартынова считает, что если в этиологии рака наследственность и играет какую-то роль, то не основную.
Что касается сходства локализации раковой опухоли, то из 4 пар ОБ, болевших раком желудочно-кишечного тракта, полное внутрипарное сходство установлено у 3 пар, т. е. у 75%. Из 22 пар РБ внутрипарное сходство найдено только у одной пары (рак желудка у обоих близнецов), т. е. в 4.5%. Это огромная разница между обоими типами близнецов. Если она будет установлена на большем материале с полной достоверностью, то толковать ее надо в том смысле, что локализация рака в большой мере обусловлена генотипом.
Внутрипарное сходство ОБ обычно не ограничивается только местом образования опухоли. Оно распространяется и на возраст появления опухоли и на гистологическую структуру ее. Пара ОБ не бывает частично похожа, только по локализации, например, или только по структуре опухоли, тогда как у РБ внутрипарное сходство ограничивается только наличием опухоли у обоих близнецов, с различием в ряде других моментов. Поразительное сходство заболевания раком ОБ отмечали и другие исследователи (ср. Gedda, 1951). Таким образом, если в возникновении рака генотип играет, по-видимому, незначительную роль, то на течение заболевания, локализацию и строение опухоли и т. д. генотип оказывает весьма существенное влияние.
Болезни близнецов в онтогенезе
В большинстве работ относительно болезней у близнецов материал рассматривается обычно на данном этапе состояния партнеров, а в случае повторного обследования тех же пар — на протяжении сравнительно короткого отрезка времени (нескольких месяцев, редко больше). Обобщения и выводы относительно заболеваний, сделанные на основании таких наблюдений, по-видимому, потребуют в ряде случаев пересмотра, переоценки и дополнений, если обследование тех же пар близнецов повторять через сравнительно значительные промежутки времени — 10 лет и больше. Такое систематическое исследование на значительном материале впервые предпринял Фершюр (Verschuer, 1954), повторно изучивший тех же близнецов через 25 лет (см. главу тринадцатую). Результат получился неожиданный для автора исследования: по всем болезням, в том числе по считавшимся автором в большой мере обусловленными наследственностью, как например туберкулез или шизофрения, однояйцевые близнецы оказались в подавляющем числе случаев дискордантными. Под руководством Фершюра аналогичное исследование было сделано его учеником Дуисом (Duis, 1956) в другом районе Германии (Франкфурт-на-Майне), правда с более коротким интервалом — через 10 лет и больше (первое обследование в 1936—1941 гг., второе — в 1951—1952 гг.). Повторно удалось обследовать 102 пары ОБ и 133 пары РБ, причем оба близнеца каждой пары были налицо. И в этом исследовании отчетливо выступила подавляющая дискордантность ОБ. Если при однократном обследовании оценка дается по схеме: здоров—болен, то при рассмотрении состояния близнецов за длительный промежуток времени выступает динамика развития заболевания и восстановления здоровья. Дуис рассматривает отдельно различные болезни и дает соответствующие таблицы вместе с описанием казуистики, а в конце приводит сводную обобщенную таблицу, в которой его данные сопоставляются с данными работы Фершюра 1954 г. Свой материал по месту сбора он обозначает «Франкфурт», а материал Фершюра — «Тюбинген». Отношение числа конкордантных пар к дискордантным дается в виде двух цифр, причем первая — это число конкордантных пар, вторая — дискордантных (табл. 44). Эти цифры дают материал для размышления медикам разных специальностей, а также генетикам. Основной вывод из них, сделанный Дуисом, — преобладающая дискордантность ОБ, по-видимому, ведет к предположению, что во всех этих заболеваниях нет не только «фатальной» предрасположенности к ним, но даже сколько-нибудь значительной роли генотипических факторов, а решающую роль играют условия жизни и тесно с ними связанное индивидуальное развитие субъекта.
Правда, если сравнить конкретные цифры конкордантных случаев с дискордантными, то можно увидеть, что в группе РБ число конкордантных случаев по отношению к числу дискордантных несколько меньше, чем в группе ОБ. Например, для морфологических аномалий вообще в группе ОБ на 19 конкордантных пар имелось 22 дискордантных (1 : 1.16), а в группе РБ — на 6 конкордантных 32 дискордантных пары (1 : 5.33). То же приблизительно вытекает из цифр по другим болезням. Это, вероятно, надо понимать в том смысле, что в случае заболевания резистентность больного, реакция его на болезнь у ОБ чаще бывает внутрипарно больше похожа, чему РБ, и это стоит в зависимости от генотипа. Но внутрипарное сходство ОБ обнаруживается в общем довольно слабо, что и можно толковать в том смысле, что наследственность в этиологии и протекании всех этих болезней имеет второстепенное значение.
Эти цифры дают материал для размышления медикам разных специальностей, а также генетикам. Основной вывод из них, сделанный Дуисом, — преобладающая дискордантность ОБ, по-видимому, ведет к предположению, что во всех этих заболеваниях нет не только «фатальной» предрасположенности к ним, но даже сколько-нибудь значительной роли генотипических факторов, а решающую роль играют условия жизни и тесно с ними связанное индивидуальное развитие субъекта.
Правда, если сравнить конкретные цифры конкордантных случаев с дискордантными, то можно увидеть, что в группе РБ число конкордантных случаев по отношению к числу дискордантных несколько меньше, чем в группе ОБ. Например, для морфологических аномалий вообще в группе ОБ на 19 конкордантных пар имелось 22 дискордантных (1 : 1.16), а в группе РБ — на 6 конкордантных 32 дискордантных пары (1 : 5.33). То же приблизительно вытекает из цифр по другим болезням. Это, вероятно, надо понимать в том смысле, что в случае заболевания резистентность больного, реакция его на болезнь у ОБ чаще бывает внутрипарно больше похожа, чему РБ, и это стоит в зависимости от генотипа. Но внутрипарное сходство ОБ обнаруживается в общем довольно слабо, что и можно толковать в том смысле, что наследственность в этиологии и протекании всех этих болезней имеет второстепенное значение.
Глава тринадцатая ВОПРОСЫ СХОДСТВА И РАЗЛИЧИЯ БЛИЗНЕЦОВ В ТЕЧЕНИЕ ЖИЗНИ
До сих пор мы имели дело с материалом, в котором сравнивались близнецы только в определенный момент их жизни или в короткий период ее. Мы рассматривали близнецов, так сказать, в статике, их жизнь бралась как бы в разрезе на определенном этапе ее. Теперь нам надо пытаться подойти к рассмотрению ряда свойств организма близнецов в динамике, в становлении, т. е. во времени, в перспективе возрастных изменений. Взаимоотношение наследственности и среды при таком подходе обнаруживается глубже и вернее. Разумеется, подобного рода исследования более трудоемки и длительны, их пока немного, а имеющиеся данные носят фрагментарный характер, далеко не давая сколько-нибудь последовательной и полной картины развития различных признаков близнецов в онтогенезе. Вес — один из тех признаков, который легко определить при рождении близнецов, так же как и рост. Как известно, вес зависит от ряда конкретных условий утробной жизни каждого из близнецов и бывает иногда очень различным у партнеров одной пары. Поэтому трудно ответить на вопрос о том, у какого из типов близнецов внутрипарная разность по весу больше, у ОБ или РБ? Разные авторы приходят к противоречивым выводам (Босик, 1934; Gedda, 1951, и др.). Средний вес близнецов в общем меньше чем вес одиночек, причем средний вес РБ немного больше, чем ОБ (Essen-Moller, 1930; Босик, 1934, и др.). Вес близнецов, по-видимому, начинает отставать от веса одиночек только после седьмого месяца беременности (Zazzo, 1955). После рождения отставание в весе близнецов постепенно ликвидируется, и на втором году жизни близнецов он достигает веса одиночек (рис. 173; Brauns, 1934, и др.). Очень мало длительных наблюдений за развитием отдельных пар близнецов. Одной из первых попыток такого рода было исследование одной пары ОБ женского пола Гизеллом с сотрудниками (Gessell a. Thompson, 1941, и др.). Об этой работе речь еще будет в дальнейшем (см. далее). Здесь же мы приведем только кривые динамики их веса и роста с первого года жизни до 12 лет (рис. 174).
Очень мало длительных наблюдений за развитием отдельных пар близнецов. Одной из первых попыток такого рода было исследование одной пары ОБ женского пола Гизеллом с сотрудниками (Gessell a. Thompson, 1941, и др.). Об этой работе речь еще будет в дальнейшем (см. далее). Здесь же мы приведем только кривые динамики их веса и роста с первого года жизни до 12 лет (рис. 174).
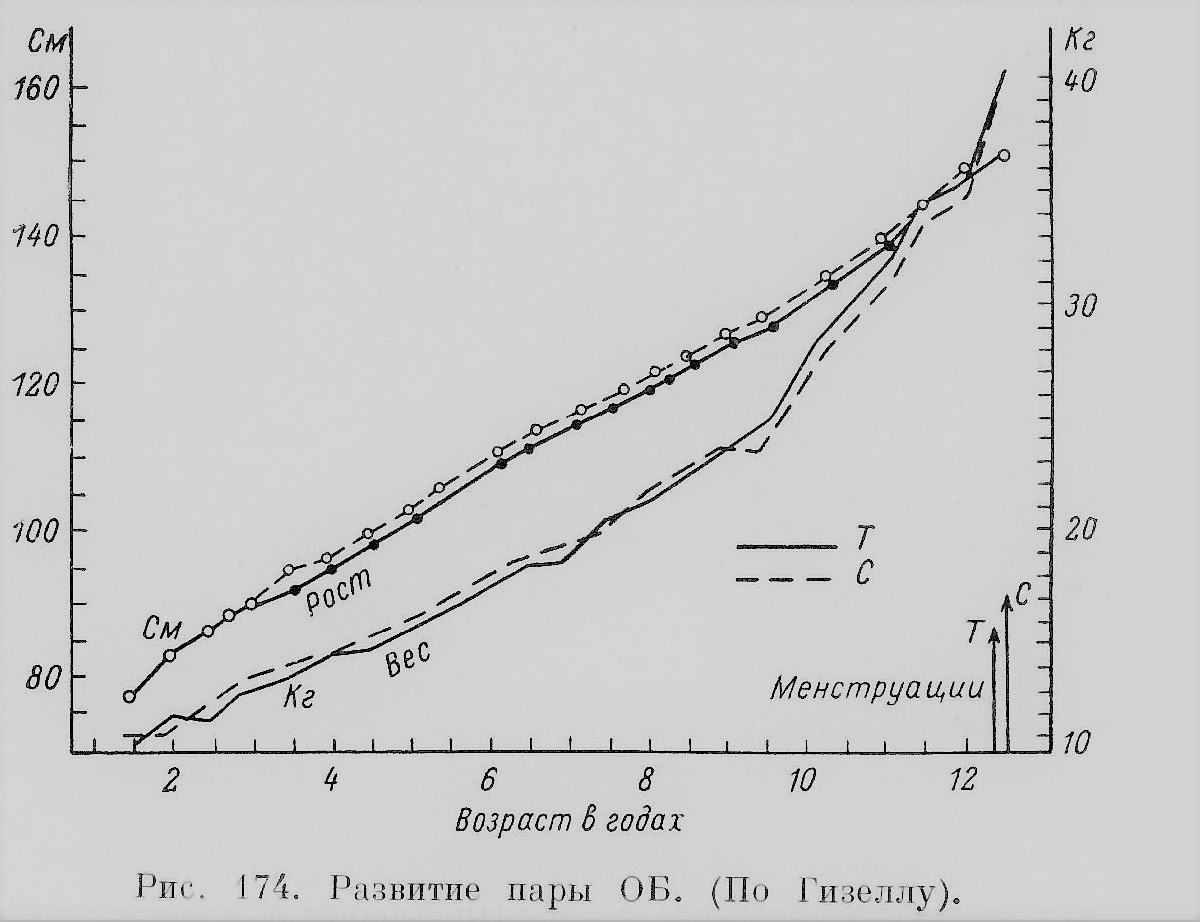
 Сходство кривых большое. Аналогичные кривые опубликованы для известной однояйцевой пятерни Дионн (рис. 175). Различие в весе и росте между этими детьми, наблюдавшееся при рождении и вызванное различием факторов утробной, среды, уже в конце первого года их жизни резко пошло на убыль, поскольку дети были поставлены в максимально благоприятные условия жизни. Сказалось их наследственное сходство, проявлению которого ничего в окружающей среде не препятствовало. Подробнее о пятерне Дионн речь идет в другом месте (см. главу девятую).
Высказывалось мнение, что отставание в весе сопровождается более легкой заболеваемостью соответствующего близнеца и т. п. (Brander, 1938), что не лишено оснований. Менее вероятно, что порядок рождения близнецов влияет на их дальнейшую судьбу, и что второй оказывается в более невыгодном положении в смысле развития, чем первый (Siebenthal, 1945; Gedda,1951).
Как в зависимости от возраста протекает развитие близнецов, наблюдаемое на таких измеримых признаках, как вес, рост, длина конечностей, объем груди, промеры головы и т. д.? Фершюр (Verschuer, 1931 —1934) сделал попытку изучить этот вопрос не путем длительных наблюдений одних и тех же пар близнецов, а путем обмера большого числа близнецов разных возрастов. Он использовал 1176 пар близнецов ОБ и РБ, взяв довольно разнородный материал (немцев, шведов и англичан), частично заимствованный из работ других ученых.
Сходство кривых большое. Аналогичные кривые опубликованы для известной однояйцевой пятерни Дионн (рис. 175). Различие в весе и росте между этими детьми, наблюдавшееся при рождении и вызванное различием факторов утробной, среды, уже в конце первого года их жизни резко пошло на убыль, поскольку дети были поставлены в максимально благоприятные условия жизни. Сказалось их наследственное сходство, проявлению которого ничего в окружающей среде не препятствовало. Подробнее о пятерне Дионн речь идет в другом месте (см. главу девятую).
Высказывалось мнение, что отставание в весе сопровождается более легкой заболеваемостью соответствующего близнеца и т. п. (Brander, 1938), что не лишено оснований. Менее вероятно, что порядок рождения близнецов влияет на их дальнейшую судьбу, и что второй оказывается в более невыгодном положении в смысле развития, чем первый (Siebenthal, 1945; Gedda,1951).
Как в зависимости от возраста протекает развитие близнецов, наблюдаемое на таких измеримых признаках, как вес, рост, длина конечностей, объем груди, промеры головы и т. д.? Фершюр (Verschuer, 1931 —1934) сделал попытку изучить этот вопрос не путем длительных наблюдений одних и тех же пар близнецов, а путем обмера большого числа близнецов разных возрастов. Он использовал 1176 пар близнецов ОБ и РБ, взяв довольно разнородный материал (немцев, шведов и англичан), частично заимствованный из работ других ученых.
 Была вычислена внутрипарная разность с помощью процентного уклонения (см. главу девятую) для ряда возрастов от первого года жизни до взрослого возраста и построены соответствующие кривые, часть которых мы здесь рассмотрим (рис. 176). Сдвиг кривых вправо показывает увеличение внутрипарной разницы. Наиболее изменчив оказался вес и больше всего у близнецов разного пола (РБр). Интересно, что разница в весе в первые годы жизни убывает. По-видимому, происходит выравнивание той разницы, которая была вызвана различием условий утробной жизни. Далее, примерно после 5 лет, начинают сказываться различия и условий среды у всех близнецов, а у РБ и генотипа. После 9 лет внутрипарная разница особенно резко выступает в связи с периодом половой зрелости и после 15 лет уменьшается: кривые образуют характерный зубец («зубец половой зрелости»), который особенно велик у РБр и продолжает далее нарастать. После 18 лет виутрнпарная разница по весу увеличивается, вероятно, в связи с увеличением различий факторов среды. Непонятно, почему в этом же возрасте виутрнпарная разница РБо идет несколько на убыль. Для объяснения нужно было бы исследовать конкретные условия жизни этой группы близнецов, чего Фершюр сделать не мог.
Внутрипарная разница в росте у всех близнецов меньше, чем в весе. Первоначальная разница у ОБ, очевидно, вызвана условиями утробной жизни. Уже с первого года жизни разница в росте у РБ заметно возрастает, а у ОБ сначала не меняется, а потом идет на убыль. В этом, очевидно, сказывается наследственная обусловленность роста, большая, чем веса. Похожую в общем картину мы видим, рассматривая кривые длины рук, промеров головы и т. д. Кривые объема груди более изменчивы и несколько напоминают кривые веса. Относительно промеров головы надо отметить заметную убыль внутрипарной разницы у ОБ, у которых эта разница, по-видимому, вызывается условиями утробной жизни и после рождения в первые годы заметно уменьшается. В некоторых случаях различие в форме головы у ОБ в детском возрасте очень заметно (рис. 34).
Те же вопросы динамики развития, темпов его надо было бы изучить на достаточно большом числе пар, наблюдаемых в течение жизни. Это дало бы возможность глубже понять ряд причин, от которых зависят внутрипарные различия близнецов на разных этапах онтогенеза по этим и другим признакам. Первую попытку в этом направлении предпринял тот же Фершюр в недавнее время. О ней речь будет дальше («Повторное обследование близнецов через 25 лет»).
Процесс окостенения скелета, тесно связанный с эндокринным аппаратом, еще мало изучен у близнецов. Об интересном исследовании этого явления на одной тройне речь была выше (см. главу десятую «Скелет»).
Появление зубов у близнецов изучалось неоднократно со времен Гальтона (Galton, 1875), который отметил появление первого зуба у двух пар близнецов в один и тот же день у каждой пары. Данные относительно начала прорезания зубов, собранные некоторыми исследователями, сведены в табл. 45.
Была вычислена внутрипарная разность с помощью процентного уклонения (см. главу девятую) для ряда возрастов от первого года жизни до взрослого возраста и построены соответствующие кривые, часть которых мы здесь рассмотрим (рис. 176). Сдвиг кривых вправо показывает увеличение внутрипарной разницы. Наиболее изменчив оказался вес и больше всего у близнецов разного пола (РБр). Интересно, что разница в весе в первые годы жизни убывает. По-видимому, происходит выравнивание той разницы, которая была вызвана различием условий утробной жизни. Далее, примерно после 5 лет, начинают сказываться различия и условий среды у всех близнецов, а у РБ и генотипа. После 9 лет внутрипарная разница особенно резко выступает в связи с периодом половой зрелости и после 15 лет уменьшается: кривые образуют характерный зубец («зубец половой зрелости»), который особенно велик у РБр и продолжает далее нарастать. После 18 лет виутрнпарная разница по весу увеличивается, вероятно, в связи с увеличением различий факторов среды. Непонятно, почему в этом же возрасте виутрнпарная разница РБо идет несколько на убыль. Для объяснения нужно было бы исследовать конкретные условия жизни этой группы близнецов, чего Фершюр сделать не мог.
Внутрипарная разница в росте у всех близнецов меньше, чем в весе. Первоначальная разница у ОБ, очевидно, вызвана условиями утробной жизни. Уже с первого года жизни разница в росте у РБ заметно возрастает, а у ОБ сначала не меняется, а потом идет на убыль. В этом, очевидно, сказывается наследственная обусловленность роста, большая, чем веса. Похожую в общем картину мы видим, рассматривая кривые длины рук, промеров головы и т. д. Кривые объема груди более изменчивы и несколько напоминают кривые веса. Относительно промеров головы надо отметить заметную убыль внутрипарной разницы у ОБ, у которых эта разница, по-видимому, вызывается условиями утробной жизни и после рождения в первые годы заметно уменьшается. В некоторых случаях различие в форме головы у ОБ в детском возрасте очень заметно (рис. 34).
Те же вопросы динамики развития, темпов его надо было бы изучить на достаточно большом числе пар, наблюдаемых в течение жизни. Это дало бы возможность глубже понять ряд причин, от которых зависят внутрипарные различия близнецов на разных этапах онтогенеза по этим и другим признакам. Первую попытку в этом направлении предпринял тот же Фершюр в недавнее время. О ней речь будет дальше («Повторное обследование близнецов через 25 лет»).
Процесс окостенения скелета, тесно связанный с эндокринным аппаратом, еще мало изучен у близнецов. Об интересном исследовании этого явления на одной тройне речь была выше (см. главу десятую «Скелет»).
Появление зубов у близнецов изучалось неоднократно со времен Гальтона (Galton, 1875), который отметил появление первого зуба у двух пар близнецов в один и тот же день у каждой пары. Данные относительно начала прорезания зубов, собранные некоторыми исследователями, сведены в табл. 45.
 Конкордантность ОБ значительно превышает таковую РБ.
Аналогичные данные получены и другими исследователями (Соболева, 1926; Korkhaus, 1929; Лихтенштейн и Басина, 1936б, и др.). Отмечалось, что сведения относительно появления первых зубов не всегда бывают очень точны, так как получаются они обычно от матерей и других родственников близнецов по воспоминаниям этих лиц. Точнее данные о «темпе» прорезания зубов, о котором можно судить непосредственно по абсолютному числу зубов у близнецов. Значительное внутрипарное сходство ОБ здесь тоже выступает очень отчетливо; оно может быть изображено в виде кривой (рис. 177).
Конкордантность ОБ значительно превышает таковую РБ.
Аналогичные данные получены и другими исследователями (Соболева, 1926; Korkhaus, 1929; Лихтенштейн и Басина, 1936б, и др.). Отмечалось, что сведения относительно появления первых зубов не всегда бывают очень точны, так как получаются они обычно от матерей и других родственников близнецов по воспоминаниям этих лиц. Точнее данные о «темпе» прорезания зубов, о котором можно судить непосредственно по абсолютному числу зубов у близнецов. Значительное внутрипарное сходство ОБ здесь тоже выступает очень отчетливо; оно может быть изображено в виде кривой (рис. 177).
 Смена молочных зубов у ОБ так же протекает с гораздо большим сходством, чем у РБ (Лихтенштейн и Басина, 1936б). О значительной роли наследственности в процессе прорезания зубов говорит также сходство его даже в деталях у отдельно изучавшихся пар (Gesell a. Thompson, 1941, и др.), а также у пятерни Дионн, у которых особенности порядка появления зубов и некоторые аномалии имели любопытное сходство (Ford a. Mason, 1943). В редких случаях наблюдалось исключительно раннее появление зубов у обоих ОБ данной пары, например при рождении у одной пары оказалось по 2 верхних резца (Newman a. Quisenberry, 1944), а у другой — по одному нижнему резцу, причем у одного близнеца это был левый резец, а у другого правый, т. е. наблюдалось явление зеркальности (Gedda, 1951). У одной пары процесс прорезания нескольких зубов шел зеркально (Steinmann, 1943).
Развитие зубной системы, ее аномалии и заболевания, легко наблюдаемые, являются удобным объектом для изучения взаимодействия наследственности с различными факторами среды и их влияний на зубы через организм близнецов (рахит, инфекции и т. п.). О болезнях зубов у двоен, различных дефектах прикуса и т. п. речь идет в другом месте (см. главу девятую «Органы пищеварения»).
Переходим к другим признакам.
Сидеть в одинаковые сроки чаще начинают партнеры ОБ, чем РБ. Так, для группы московских близнецов, состоящей из 164 пар, коэффициент корреляции для ОБ оказался очень большим — 0.97, для РБ — 0.73 (Босяк, 1934).
Начало хождения менее конкордантно у ОБ, чем начало сидения. Из 257 пар московских близнецов 67.0% ОБ начали ходить одновременно и 17.5% с разницей до одного месяца; для РБо соответствующие цифры оказались 29.9% и 29.9%, а для РБр — 44.4% и 20.6%. Если среднюю внутрипарную разницу начала хождения выразить в коэффициентах корреляции, то для ОБ такой коэффициент будет 0.89, для РБо 0.74, а для РБр — 0.58. Несколько меньшая конкордантность ОБ найдена на другом материале (Brauns, 1934; Kruger, 1936. и др.).
В течение первого года жизни близнецы в массе будто бы отстают в развитии от одиночек (Соболева, 1926). Это, по-видимому, наблюдается даже у близнецов, родившихся вовремя а не только у недоносков. Так, у 55 пар близнецов обоих типов, своевременно родившихся, установлено отставание в психомоторике, изучавшейся с помощью лестницы Брюне. Отставание заметнее у ОБ и вообще у мальчиков. Очень существенно здесь влияние социально-экономических факторов (Lezine, 1953).
Начало речи — более сложный вопрос. Социальная среда, влияние окружающих, одного близнеца на другого — все это имеет очень большое значение для различных сторон развития речи. Этот вопрос еще пока недостаточно изучен (ср. Davis, 1941). Имеются указания, что близнецы обоих типов начинают говорить более или менее одновременно, что внутрипарная разница у ОБ исчезает в течение первого года, а у РБ, наоборот, возрастает (Gedda, 1951). Разные пары близнецов, разумеется, начинают говорить в разное время, но в среднем близнецы опаздывают на 4—8 месяцев по сравнению с одиночками (Schwesinger, 1940). Так, например, по числу слов, произношению и проч. близнецы в среднем отстают от одиночек (Day,1932), как это видно из приводимого графика (рис. 178).
Смена молочных зубов у ОБ так же протекает с гораздо большим сходством, чем у РБ (Лихтенштейн и Басина, 1936б). О значительной роли наследственности в процессе прорезания зубов говорит также сходство его даже в деталях у отдельно изучавшихся пар (Gesell a. Thompson, 1941, и др.), а также у пятерни Дионн, у которых особенности порядка появления зубов и некоторые аномалии имели любопытное сходство (Ford a. Mason, 1943). В редких случаях наблюдалось исключительно раннее появление зубов у обоих ОБ данной пары, например при рождении у одной пары оказалось по 2 верхних резца (Newman a. Quisenberry, 1944), а у другой — по одному нижнему резцу, причем у одного близнеца это был левый резец, а у другого правый, т. е. наблюдалось явление зеркальности (Gedda, 1951). У одной пары процесс прорезания нескольких зубов шел зеркально (Steinmann, 1943).
Развитие зубной системы, ее аномалии и заболевания, легко наблюдаемые, являются удобным объектом для изучения взаимодействия наследственности с различными факторами среды и их влияний на зубы через организм близнецов (рахит, инфекции и т. п.). О болезнях зубов у двоен, различных дефектах прикуса и т. п. речь идет в другом месте (см. главу девятую «Органы пищеварения»).
Переходим к другим признакам.
Сидеть в одинаковые сроки чаще начинают партнеры ОБ, чем РБ. Так, для группы московских близнецов, состоящей из 164 пар, коэффициент корреляции для ОБ оказался очень большим — 0.97, для РБ — 0.73 (Босяк, 1934).
Начало хождения менее конкордантно у ОБ, чем начало сидения. Из 257 пар московских близнецов 67.0% ОБ начали ходить одновременно и 17.5% с разницей до одного месяца; для РБо соответствующие цифры оказались 29.9% и 29.9%, а для РБр — 44.4% и 20.6%. Если среднюю внутрипарную разницу начала хождения выразить в коэффициентах корреляции, то для ОБ такой коэффициент будет 0.89, для РБо 0.74, а для РБр — 0.58. Несколько меньшая конкордантность ОБ найдена на другом материале (Brauns, 1934; Kruger, 1936. и др.).
В течение первого года жизни близнецы в массе будто бы отстают в развитии от одиночек (Соболева, 1926). Это, по-видимому, наблюдается даже у близнецов, родившихся вовремя а не только у недоносков. Так, у 55 пар близнецов обоих типов, своевременно родившихся, установлено отставание в психомоторике, изучавшейся с помощью лестницы Брюне. Отставание заметнее у ОБ и вообще у мальчиков. Очень существенно здесь влияние социально-экономических факторов (Lezine, 1953).
Начало речи — более сложный вопрос. Социальная среда, влияние окружающих, одного близнеца на другого — все это имеет очень большое значение для различных сторон развития речи. Этот вопрос еще пока недостаточно изучен (ср. Davis, 1941). Имеются указания, что близнецы обоих типов начинают говорить более или менее одновременно, что внутрипарная разница у ОБ исчезает в течение первого года, а у РБ, наоборот, возрастает (Gedda, 1951). Разные пары близнецов, разумеется, начинают говорить в разное время, но в среднем близнецы опаздывают на 4—8 месяцев по сравнению с одиночками (Schwesinger, 1940). Так, например, по числу слов, произношению и проч. близнецы в среднем отстают от одиночек (Day,1932), как это видно из приводимого графика (рис. 178).
 Разумеется, не каждая пара близнецов отстает в развитии речи. Браунс (Brauns, 1934) приводит конкретные данные о сроках начала речи близнецов, правда полученные со слов родителей, что ставит, конечно, под сомнение точность таких данных. Так, 40 человек стали говорить на первом году, 16 — к 15 месяцам, 35 — к 13/4 года; 24 ребенка научились говорить за период 1 3/4—2 1/4 года, 7 — к 1 3/4 годам и 10 — после 3 лет. По другим данным, из 28 пар ОБ у 18 речь развивалась нормально, у 7 с опозданием (в период от 18 месяцев до 2 лет), а в 3 случаях с явной задержкой — до 3-го и 4-го года (Luchsinger, 1940). Причина такой задержки не ясна. У мальчиков отставание встречается чаще, чем у девочек. Вряд ли это можно объяснить одной наследственностью (Seemann, 1937). Необходимо дальнейшее изучение этого вопроса. Возможно, что в некоторых случаях отставание развития речи у близнецов зависит от известной замкнутости их как пары относительно окружающего мира, как например у пятерни Дионн или пары ОБ, изученной Лурией и Юдович (1956). В качестве крайнего примера могут служить такие редкие случаи, когда у пары близнецов возникал своеобразный собственный язык, непонятный для посторонних. Так, например, пара незаконнорожденных близнецов в возрасте около 4 лет, покинутых матерью и живших изолированно в одной датской деревне с немой старухой, бойко объяснялись на каком-то непонятном для других языке, хотя большинство «слов» этого языка напоминало датский язык; настоящий датский язык эти близнецы не знали (Jespersen, 1925). Близнецы Дионн тоже объяснялись между собой на каком-то своеобразном детском языке, а вообще в развитии речи отставали от нормы (Schwesinger, 1940).
Недавно были проведены новые наблюдения над развитием речи и также некоторых дефектов ее (заикания и др.) у близнецов (Luchsinger, 1953). Из 12 пар ОБ, из которых каждая пара жила примерно в одинаковых условиях, 10 пар имели нормальное развитие речи, а 2 пары — запаздывающее (первые слова около 2 1/2 и 4 лет). Внутрипарное сходство ОБ по скорости развития речи было велико. Наоборот, среди 12 пар РБ внутрипарная дискордантность в общем была значительно больше. Автор не мог проанализировать этот сложный процесс. Эта задача будущих исследователей.
Наступление половой зрелости у близнецов еще мало изучено. На сравнительно небольшом материале установлено, что изменение голоса у ОБ мужского пола происходит с большим внутрипарным сходством (Luchsinger, 1944). Другие стороны полового созревания мальчиков-близнецов систематически не изучены.
Разумеется, не каждая пара близнецов отстает в развитии речи. Браунс (Brauns, 1934) приводит конкретные данные о сроках начала речи близнецов, правда полученные со слов родителей, что ставит, конечно, под сомнение точность таких данных. Так, 40 человек стали говорить на первом году, 16 — к 15 месяцам, 35 — к 13/4 года; 24 ребенка научились говорить за период 1 3/4—2 1/4 года, 7 — к 1 3/4 годам и 10 — после 3 лет. По другим данным, из 28 пар ОБ у 18 речь развивалась нормально, у 7 с опозданием (в период от 18 месяцев до 2 лет), а в 3 случаях с явной задержкой — до 3-го и 4-го года (Luchsinger, 1940). Причина такой задержки не ясна. У мальчиков отставание встречается чаще, чем у девочек. Вряд ли это можно объяснить одной наследственностью (Seemann, 1937). Необходимо дальнейшее изучение этого вопроса. Возможно, что в некоторых случаях отставание развития речи у близнецов зависит от известной замкнутости их как пары относительно окружающего мира, как например у пятерни Дионн или пары ОБ, изученной Лурией и Юдович (1956). В качестве крайнего примера могут служить такие редкие случаи, когда у пары близнецов возникал своеобразный собственный язык, непонятный для посторонних. Так, например, пара незаконнорожденных близнецов в возрасте около 4 лет, покинутых матерью и живших изолированно в одной датской деревне с немой старухой, бойко объяснялись на каком-то непонятном для других языке, хотя большинство «слов» этого языка напоминало датский язык; настоящий датский язык эти близнецы не знали (Jespersen, 1925). Близнецы Дионн тоже объяснялись между собой на каком-то своеобразном детском языке, а вообще в развитии речи отставали от нормы (Schwesinger, 1940).
Недавно были проведены новые наблюдения над развитием речи и также некоторых дефектов ее (заикания и др.) у близнецов (Luchsinger, 1953). Из 12 пар ОБ, из которых каждая пара жила примерно в одинаковых условиях, 10 пар имели нормальное развитие речи, а 2 пары — запаздывающее (первые слова около 2 1/2 и 4 лет). Внутрипарное сходство ОБ по скорости развития речи было велико. Наоборот, среди 12 пар РБ внутрипарная дискордантность в общем была значительно больше. Автор не мог проанализировать этот сложный процесс. Эта задача будущих исследователей.
Наступление половой зрелости у близнецов еще мало изучено. На сравнительно небольшом материале установлено, что изменение голоса у ОБ мужского пола происходит с большим внутрипарным сходством (Luchsinger, 1944). Другие стороны полового созревания мальчиков-близнецов систематически не изучены.
 Созревание девочек-близнецов изучалось по первой менструации (Petri, 1934; Feigel, 1936, и др.). Внутрипарное сходство ОБ в этом отношении при сравнении с РБ, родственницами и популяцией бросается в глаза (рис. 179). Из пары ОБ обычно первая менструация начинается раньше у той близнячки, которая при рождении была менее тяжелой и менее крепкой (Turpin et Tisserand, 1938). Развитие вторичных половых признаков у близнецов женского пола еще мало изучено. Есть данные, что груди у ОБ развиваются конкордантно, а у РБ этого не наблюдается (Birkenfeld, 1932, и др.).
Дальнейшее развитие половой жизни близнецов чрезвычайно осложняется социально-экономическими факторами и потому ускользает от систематического исследования.
Не раз возникал вопрос, в какой мере процесс старения и продолжительность жизни зависит от генотипа. Еще со времени Гальтона были попытки показать на близнецах, что срок жизни человека предопределяется наследственностью (Curtius u. Korkhaus, 1930; Vogl et al., 1939, и др.). За последнее время опубликованы данные, которые показывают несостоятельность такого предположения. Речь о них будет ниже. Однако сравнительно недавно была сделана еще одна попытка найти факты в пользу этой идеи. Я имею в виду крупное по объему исследование Коллмана и Сандера (Kallmann а. Sander, 1948, 1949). В указанных статьях приводятся итоги первых 3 лет работы с охватом 933 пар близнецов в возрасте свыше 60 лет, жителей Нью-Порка и его окрестностей. У 431 пары были живы оба близнеца. Из общего числа всех изучавшихся близнецов 697 человек было мужского пола и 905 женского, т. е. число женщин этого возраста было заметно больше, что в общем соответствует обычному возрастному соотношению полов в населении. В собранном материале число пар ОБ составляло около 30% всех близнецов, и авторы считают, что это говорит о безотборности их серии близнецов (см. главу десятую). Сходство некоторых пар ОБ на протяжении всех возрастов очень велико, примером чего служит приведенная серия фотографий (рис. 180).
Созревание девочек-близнецов изучалось по первой менструации (Petri, 1934; Feigel, 1936, и др.). Внутрипарное сходство ОБ в этом отношении при сравнении с РБ, родственницами и популяцией бросается в глаза (рис. 179). Из пары ОБ обычно первая менструация начинается раньше у той близнячки, которая при рождении была менее тяжелой и менее крепкой (Turpin et Tisserand, 1938). Развитие вторичных половых признаков у близнецов женского пола еще мало изучено. Есть данные, что груди у ОБ развиваются конкордантно, а у РБ этого не наблюдается (Birkenfeld, 1932, и др.).
Дальнейшее развитие половой жизни близнецов чрезвычайно осложняется социально-экономическими факторами и потому ускользает от систематического исследования.
Не раз возникал вопрос, в какой мере процесс старения и продолжительность жизни зависит от генотипа. Еще со времени Гальтона были попытки показать на близнецах, что срок жизни человека предопределяется наследственностью (Curtius u. Korkhaus, 1930; Vogl et al., 1939, и др.). За последнее время опубликованы данные, которые показывают несостоятельность такого предположения. Речь о них будет ниже. Однако сравнительно недавно была сделана еще одна попытка найти факты в пользу этой идеи. Я имею в виду крупное по объему исследование Коллмана и Сандера (Kallmann а. Sander, 1948, 1949). В указанных статьях приводятся итоги первых 3 лет работы с охватом 933 пар близнецов в возрасте свыше 60 лет, жителей Нью-Порка и его окрестностей. У 431 пары были живы оба близнеца. Из общего числа всех изучавшихся близнецов 697 человек было мужского пола и 905 женского, т. е. число женщин этого возраста было заметно больше, что в общем соответствует обычному возрастному соотношению полов в населении. В собранном материале число пар ОБ составляло около 30% всех близнецов, и авторы считают, что это говорит о безотборности их серии близнецов (см. главу десятую). Сходство некоторых пар ОБ на протяжении всех возрастов очень велико, примером чего служит приведенная серия фотографий (рис. 180).


 Симптомы старости, например облысение, развитие старческого психоза и т. п., у некоторых пар установлены очень похожие, даже несмотря на известные различия в условиях жизни и биографии. Внутрипарные различия психики у стариков, установленные с помощью различных тестов, оказались у ОБ в общем меньше, чем у РБ. Ряд аналогичных данных сообщался и раньше (рис. 181, 182; ср.: Lotze, 1937, и др.).
Что касается долголетия, то оно оценивалось путем вычисления средней разницы в месяцах между смертью одного из близнецов и другого той же пары, умерших по «естественным» причинам после 60 лет. Таких пар было в серии 58. Для группы ОБ эта средняя равнялась 36.9 месяца, для РБо — 78.3, для РБр — 126.6 месяца. Авторы видят в этих цифрах свидетельство в пользу существенной роли генотипа для продолжительности жизни. Однако Фершюр (Verschuer, 1954) все же считает этот материал американцев отобранным на долголетие и с их выводами не согласен. Он противопоставляет им свой материал, собранный без всякого отбора и состоящий из немецких близнецов разного возраста. Эти данные сведены в табл. 46.
Симптомы старости, например облысение, развитие старческого психоза и т. п., у некоторых пар установлены очень похожие, даже несмотря на известные различия в условиях жизни и биографии. Внутрипарные различия психики у стариков, установленные с помощью различных тестов, оказались у ОБ в общем меньше, чем у РБ. Ряд аналогичных данных сообщался и раньше (рис. 181, 182; ср.: Lotze, 1937, и др.).
Что касается долголетия, то оно оценивалось путем вычисления средней разницы в месяцах между смертью одного из близнецов и другого той же пары, умерших по «естественным» причинам после 60 лет. Таких пар было в серии 58. Для группы ОБ эта средняя равнялась 36.9 месяца, для РБо — 78.3, для РБр — 126.6 месяца. Авторы видят в этих цифрах свидетельство в пользу существенной роли генотипа для продолжительности жизни. Однако Фершюр (Verschuer, 1954) все же считает этот материал американцев отобранным на долголетие и с их выводами не согласен. Он противопоставляет им свой материал, собранный без всякого отбора и состоящий из немецких близнецов разного возраста. Эти данные сведены в табл. 46.
 Из этих цифр видно, что большей конкордантности между ОБ по сравнению с РБ в смысле долголетия нет. Так. в случае смерти обоих близнецов женского пола средний промежуток между смертями для ОБ 8 лет 10 мес., а для РБ — 6 лет и 4 мес. Также и в случае смерти одного из близнецов-женщин оставшаяся в живых в среднем дольше жила из группы ОБ, а не РБ, и т. д.
Рассмотрение конкретных причин смерти близнецов в тех случаях, когда умерли оба, показывает, что в большинстве случаев они умерли от совсем разных причин, если не считать гибель двух близнецов в одном и том же бою во время последней войны или смерть двух 83-летних близнецов от воспаления легких через 4 месяца один после другого. Даже при наличии преимущественно наследственных болезней, таких, как например, шизофрения, от такой болезни погибал только один из ОБ. Констатируя факты продолжительной жизни одного из ОБ после смерти другого, Фершюр считает, что собранный им материал опровергает ходячий предрассудок, согласно которому смерть одного из ОБ влечет за собой смерть другого и что срок жизни их предопределен наследственностью, как это еще недавно пытались показать на подобранном материале (Bruins, 1955). Таким образом, разговоры о «фатальной» наследственности в отношении продолжительности жизни, очевидно, оказываются несостоятельными. Однако различная степень «стойкости» организма, отчасти зависящая от генотипа, конечно, этим не отрицается. Долголетие в каждом случае зависит от многообразного соотношения условий жизни, пережитых состояний организма и его природных данных. Поскольку биография каждого из однояйцевых близнецов может слагаться порой очень различно, то и смерть каждого из них может прийти в очень разные сроки, не говоря о «случайных» причинах ее.
Недавно Коллман с сотрудниками (Kallmann et al., 1956) опубликовал новую работу о долголетии близнецов и их родственников. В этой работе имеются данные о 180 парах близнецов, достигших 60 лет, где оба близнеца каждой пары умерли. Разница в сроке смерти между членами одной пары для ОБ составляет около 36 месяцев, для РБ одинакового пола — около 74 месяцев, а для РБ разного пола — около 106 месяцев. Эти цифры близки к ранее полученным Коллманом и толкуются им как ясная демонстрация меньшей внутрипарной разницы ОБ, а, следовательно, меньшего влияния генотипа на продолжительность жизни. Он оспаривает вышеприведенные выводы Фершюра (Verschuer, 1954), считая, что они основаны на небольшом материале (18 пар близнецов) и притом немцах, живших в период гитлеровского режима, когда риск гибели был одинаково велик для всех, особенно же для мужчин среднего возраста, что стирает разницу между ОБ и РБ.
Далее Коллман с сотрудниками утверждают на основании своего материала, что ОБ внутрипарно в 2 раза более похожи, чем РБ, в смысле сходства причин смерти обоих партнеров. Наконец, на основании большого материала — 2536 человек близнецов в возрасте свыше 60 лет, их родителей и братьев-сестер, авторы приходят к выводу, что существует зависимость между долголетием родителей и детей, с полом не связанная, иначе говоря, у родителей, дольше живших, и дети в среднем жили дольше. Это авторы также толкуют как доказательство роли генотипа для долголетия. К сожалению, весь материал изложен очень кратко и абстрактно; конкретные пары, их особенности и течение их жизни не описаны, и потому в таких вопросах, как, например, причины смерти близнецов, остается неясным, какие случаи принимались авторами за случаи с одинаковыми причинами смерти и т. д. Более детальное рассмотрение материала, подобного собранному Коллманом, необходимо, так как без этого невозможно решить вопрос, в чем же конкретно выражается генотипическое предрасположение к долголетию и какие факторы среды препятствуют ему. Для суждения о роли генотипа для долголетия нужен, конечно, и близнецовый и родословный материал, собранный в других странах на различных группах населения, а не только в одном городе. Это дело будущих исследований, а работы Коллмана, как и Фершюра, являются лишь первыми шагами в исследовании вопроса генетики долголетия человека.
Взаимодействие наследственности и среды в жизни отдельных пар близнецов изучалось разными путями. Рассмотрим три из них, являющихся тремя продуктивными направлениями современной гемеллологии: 1) длительное наблюдение с привлечением эксперимента над парой близнецов с раннего детства до половой зрелости; 2) однократное обследование пары ОБ, живших значительную часть жизни врозь, порой в очень разных условиях; 3) повторное обследование тех же пар близнецов через 25 лет.
Из этих цифр видно, что большей конкордантности между ОБ по сравнению с РБ в смысле долголетия нет. Так. в случае смерти обоих близнецов женского пола средний промежуток между смертями для ОБ 8 лет 10 мес., а для РБ — 6 лет и 4 мес. Также и в случае смерти одного из близнецов-женщин оставшаяся в живых в среднем дольше жила из группы ОБ, а не РБ, и т. д.
Рассмотрение конкретных причин смерти близнецов в тех случаях, когда умерли оба, показывает, что в большинстве случаев они умерли от совсем разных причин, если не считать гибель двух близнецов в одном и том же бою во время последней войны или смерть двух 83-летних близнецов от воспаления легких через 4 месяца один после другого. Даже при наличии преимущественно наследственных болезней, таких, как например, шизофрения, от такой болезни погибал только один из ОБ. Констатируя факты продолжительной жизни одного из ОБ после смерти другого, Фершюр считает, что собранный им материал опровергает ходячий предрассудок, согласно которому смерть одного из ОБ влечет за собой смерть другого и что срок жизни их предопределен наследственностью, как это еще недавно пытались показать на подобранном материале (Bruins, 1955). Таким образом, разговоры о «фатальной» наследственности в отношении продолжительности жизни, очевидно, оказываются несостоятельными. Однако различная степень «стойкости» организма, отчасти зависящая от генотипа, конечно, этим не отрицается. Долголетие в каждом случае зависит от многообразного соотношения условий жизни, пережитых состояний организма и его природных данных. Поскольку биография каждого из однояйцевых близнецов может слагаться порой очень различно, то и смерть каждого из них может прийти в очень разные сроки, не говоря о «случайных» причинах ее.
Недавно Коллман с сотрудниками (Kallmann et al., 1956) опубликовал новую работу о долголетии близнецов и их родственников. В этой работе имеются данные о 180 парах близнецов, достигших 60 лет, где оба близнеца каждой пары умерли. Разница в сроке смерти между членами одной пары для ОБ составляет около 36 месяцев, для РБ одинакового пола — около 74 месяцев, а для РБ разного пола — около 106 месяцев. Эти цифры близки к ранее полученным Коллманом и толкуются им как ясная демонстрация меньшей внутрипарной разницы ОБ, а, следовательно, меньшего влияния генотипа на продолжительность жизни. Он оспаривает вышеприведенные выводы Фершюра (Verschuer, 1954), считая, что они основаны на небольшом материале (18 пар близнецов) и притом немцах, живших в период гитлеровского режима, когда риск гибели был одинаково велик для всех, особенно же для мужчин среднего возраста, что стирает разницу между ОБ и РБ.
Далее Коллман с сотрудниками утверждают на основании своего материала, что ОБ внутрипарно в 2 раза более похожи, чем РБ, в смысле сходства причин смерти обоих партнеров. Наконец, на основании большого материала — 2536 человек близнецов в возрасте свыше 60 лет, их родителей и братьев-сестер, авторы приходят к выводу, что существует зависимость между долголетием родителей и детей, с полом не связанная, иначе говоря, у родителей, дольше живших, и дети в среднем жили дольше. Это авторы также толкуют как доказательство роли генотипа для долголетия. К сожалению, весь материал изложен очень кратко и абстрактно; конкретные пары, их особенности и течение их жизни не описаны, и потому в таких вопросах, как, например, причины смерти близнецов, остается неясным, какие случаи принимались авторами за случаи с одинаковыми причинами смерти и т. д. Более детальное рассмотрение материала, подобного собранному Коллманом, необходимо, так как без этого невозможно решить вопрос, в чем же конкретно выражается генотипическое предрасположение к долголетию и какие факторы среды препятствуют ему. Для суждения о роли генотипа для долголетия нужен, конечно, и близнецовый и родословный материал, собранный в других странах на различных группах населения, а не только в одном городе. Это дело будущих исследований, а работы Коллмана, как и Фершюра, являются лишь первыми шагами в исследовании вопроса генетики долголетия человека.
Взаимодействие наследственности и среды в жизни отдельных пар близнецов изучалось разными путями. Рассмотрим три из них, являющихся тремя продуктивными направлениями современной гемеллологии: 1) длительное наблюдение с привлечением эксперимента над парой близнецов с раннего детства до половой зрелости; 2) однократное обследование пары ОБ, живших значительную часть жизни врозь, порой в очень разных условиях; 3) повторное обследование тех же пар близнецов через 25 лет.
Длительное изучение развития одной пары ОБ
Образцом длительного изучения одной пары ОБ была работа психолога Гизелла с сотрудниками (Strayer, 1930; Thompson, 1932; Hilgard, 1933; Gesell a. Blake, 1936; Gesell a. Thompson, 1941, 1952). Исследование началось, когда младенцам шел только первый год, а закончилось, когда им было 14 лет. Близнецы были женского пола и условно обозначались буквами Т и С, поскольку первую в ряде случаев тренировали с экспериментальной целью, а вторая служила контролем. Ход физического развития детей был очень похож, их рост и веса изображены графически на рис. 174.
При рождении близнецы были почти одинакового веса и одинаковой длины. В дальнейшем девочка С, родившаяся на полчаса раньше Т, стала немного выше своей близнячки и была почти все время тяжелее, вплоть до 10-го года. Зубы появились раньше у Т, менструации тоже. Порядок появления первых 12 постоянных зубов был у обеих одинаков. История их заболеваний также очень похожа, за исключением небольших различий, например Т оказалась более подверженной инфекциям верхних дыхательных путей и вообще медленнее выздоравливала после инфекций.
Уже вскоре после рождения было замечено, что Т более «активна», чем С. И в дальнейшем Т опережала С в отношении моторики. Ее движения энергичнее и законченнее, например в танце; она вообще подвижнее и четче в спорте и т. п. Однако в области тонких движений С превосходит Т, например в работе кисточкой или пером, в вязании крючком. Интересна картина эволюции их почерков с 7 до 14 лет. Порой их почерк неразличим, в общем же Т выводит буквы четче, чем С. У Т более выражена функциональная асимметрия, чем у С.
В интеллектуальном отношении, судя по школьным успехам и ряду тестов, Т слегка опережает С. Наблюдается ряд различии в проявлении «внимания» у них. Так, у Т внимание более интенсивно и четко фиксируется на объекте, у С более слабо и расплывчато и т. д.
В области речи у близнецов тоже установлены известные различия: С с раннего возраста была более болтлива, а вообще более общительна — дома и в школе, охотнее пишет письма и т. д. Однако Т немного лучше произносила слова, ее словарь был несколько больше, она чаще спрашивала. В общем же развитие их в области речи очень похоже, о чем говорилось в другом месте.
В детстве С больше интересовалась своей близнячкой, чем последняя ею. Хотя и в слабой степени, но С была «лидером» их пары. Среди товарищей по школе С несколько популярнее, чем Т. В детстве обе хотели быть учительницей. Но позже Т желала стать лавочницей или кормилицей, а С — принцессой или певицей.
Причины, вызвавшие индивидуальные особенности у близнецов, остаются неясными. В течение 14 лет они жили вместе в одной семье, в общем в одинаковых условиях, почти не разлучаясь. Только попав в школу, они оказались в параллельных классах, у разных учителей. Изо дня в день они находились в разном «школьном климате». Но исследователи не установили какого-либо влияния этого различия среды в пользу или во вред одной из близнячек, не обнаружили какого-либо постоянного психического расхождения из-за этого. Мало того, те сотни часов, которые Т находилась в условиях экспериментов по моторике, речи и т. п., не прибавили ничего существенного к ее индивидуальности. В известной мере такая тренировка пошла ей, конечно, на пользу, помогла, надо думать, достигнуть известных успехов в спорте и т. д., но достоверных данных в этом отношении нет. Авторы не были в состоянии установить факторы, обусловившие индивидуальные особенности близнецов Т и С.
Это трудоемкое интересное исследование Гизелла с сотрудниками является до сих пор единственным в этом роде в мировой литературе. Нечто подобное можно было бы ожидать от многочисленных исследований пятерни Дионн, которые все уже благополучно прожили свыше 20 лет, находясь под постоянным разносторонним наблюдением, особенно в первые годы жизни. Но, по-видимому, не было одного постоянного руководителя по изучению Дионн, полученный за все истекшее время материал не объединен и не опубликован в виде целого исследования; имеется лишь ряд отдельных работ, касающихся главным образом специальных вопросов; на некоторые из них мы уже ссылались.
Было бы, конечно, чрезвычайно интересно повторить исследование по методу Гизелла на нескольких парах ОБ, проводя изучение развития мозга детей не только психологическими методами, но и максимально используя условнорефлекторные методики. Изучение физиологии высшей нервной деятельности, вероятно, поможет обнаружить хотя бы некоторые факторы, вызвавшие различия «склада высшей нервной деятельности» близнецов, различия их «характеров», особенностей их индивидуальностей, что не удалось американским ученым. Описанная ими картина развития пары ОБ, при всех недостатках этой картины, является стимулирующим примером для новых исследований такого рода, ставящих себе задачей объяснить причины внутрипарных различий ОБ путем изучения становления этих различий.
ОБ, воспитанные врозь
В основе таких исследований поставлен вопрос: какие внутрипарные различия возникают у ОБ, выросших в различных условиях жизни. Это, разумеется, сложный вопрос. Трудность его становится понятнее при первых же попытках его конкретного разрешения. Несомненные различия у таких близнецов были установлены на первых же изученных парах (Muller, 1925; Wagenseil, 1931, и др.). После этого большое систематическое исследование было предпринято Ньюменом с сотрудниками — психологами и статистиком (Newman et al., 1937; Gardner a. Newman, 1940; Newman, 1940c). Авторы в меру своих возможностей тщательно изучили 20 пар ОБ, живших врозь, и сравнили их как внутрипарно, так и статистически с близнецами, жившими в «одинаковых» условиях (по 50 пар ОБ и РБ). Эти 20 пар представляют несомненно большую редкость иявляются интересным материалом. В качестве примера мы кратко рассмотрим 5 из них, взяв прежде всего 4 пары из числа живших в наиболее различных условиях. Мери и Мебл 29 лет (рис. 183) были разлучены в возрасте 5 месяцев и усыновлены в разных семьях. До 6 лет обе жили на фермах своих приемных родителей и иногда виделись. В дальнейшем Мери переселилась в небольшой город и осталась там жить, тогда как Мебл продолжала вести типичную фермерскую жизнь. Мери удалось окончить высшую школу. Она выполняла канцелярскую работу и давала по вечерам уроки музыки, которой занималась с 10 лет. Мебл окончила только сельскую школу. Читать ей пришлось мало. Обе жили в относительном достатке, примерно на одном экономическом уровне, но в разном социальном окружении. Состав семей, в которых они жили, был разный. Мебл жила среди мальчиков и девочек близкого ей возраста, Мери — среди старших и т. д. При обследовании этих близнецов в возрасте 29 лет фермерша Мебл оказалась значительно тяжелее (на 28 фунтов), с более развитой мускулатурой. Рост их был почти одинаков, однако Мебл немного выше (на 3.7 см). Размеры головы были также почти одинаковы. Психика их изучалась с помощью тестов. Умственное развитие Мери, как и следовало ожидать, было выше; она оказалась более «возбудима», а Мебл — более «флегматична», однако при этом и более агрессивна, деловита и смела. Она шагала твердой, мужественной походкой, в отличие от Мери, у которой походка и манеры были более «дамские».
Гледис и Хелен, 35 лет при обследовании. Разлучены были в возрасте около 11 месяцев и встретились только в 28 лет. Воспитывались они в разных условиях: Гледис в семье железнодорожника, Хелен на ферме, а позже в городе. Образование их было очень различным: Хелен кончила высшую школу и стала учительницей, тогда как Гледис не удалось кончить даже среднюю школу, и она работала в качестве мелкой служащей. Социальное окружение их отличалось во многих отношениях, хотя обе жили в городе. По росту, весу и т. д. у них разница невелика. Однако Хелен, жизнь которой сложилась удачнее, сохранилась физически лучше, чем ее близнячка. Различие в их умственном развитии очень заметно даже без применения тестов. Хелен, учительница, более слащава и любезна, более заботится о своей внешности и благоприятном впечатлении на окружающих. Гледис гораздо проще и мало озабочена производимым впечатлением.
Третья пара — Милфрид и Рут. Рано разлученные, они воспитывались в совсем разных семьях. Первая жила в доме банкира города средней величины, получила хорошее образование и вращалась среди интеллигентных людей, посещавших дом ее приемных родителей. Рут попала в семью малообразованных людей из рабочих. Угнетенное положение в доме отразилось на ее характере: заторможенная, пугливая, недоверчивая, молчаливая, она говорила в лепечущей манере, неудачно выражаясь. Милфрид, наоборот, была общительна, свободно и бойко говорила. И хотя обе получили высшее образование, Милфрид согласно оценке с помощью тестов, была более развита.
Эти 3 пары принадлежат к тем, которые из числа изучавшихся 20 пар имели ряд очень значительных различий в условиях жизни, в зависимости от чего можно поставить некоторые внутрипарные различия этих близнецов, особенно психики.
Мери и Мебл 29 лет (рис. 183) были разлучены в возрасте 5 месяцев и усыновлены в разных семьях. До 6 лет обе жили на фермах своих приемных родителей и иногда виделись. В дальнейшем Мери переселилась в небольшой город и осталась там жить, тогда как Мебл продолжала вести типичную фермерскую жизнь. Мери удалось окончить высшую школу. Она выполняла канцелярскую работу и давала по вечерам уроки музыки, которой занималась с 10 лет. Мебл окончила только сельскую школу. Читать ей пришлось мало. Обе жили в относительном достатке, примерно на одном экономическом уровне, но в разном социальном окружении. Состав семей, в которых они жили, был разный. Мебл жила среди мальчиков и девочек близкого ей возраста, Мери — среди старших и т. д. При обследовании этих близнецов в возрасте 29 лет фермерша Мебл оказалась значительно тяжелее (на 28 фунтов), с более развитой мускулатурой. Рост их был почти одинаков, однако Мебл немного выше (на 3.7 см). Размеры головы были также почти одинаковы. Психика их изучалась с помощью тестов. Умственное развитие Мери, как и следовало ожидать, было выше; она оказалась более «возбудима», а Мебл — более «флегматична», однако при этом и более агрессивна, деловита и смела. Она шагала твердой, мужественной походкой, в отличие от Мери, у которой походка и манеры были более «дамские».
Гледис и Хелен, 35 лет при обследовании. Разлучены были в возрасте около 11 месяцев и встретились только в 28 лет. Воспитывались они в разных условиях: Гледис в семье железнодорожника, Хелен на ферме, а позже в городе. Образование их было очень различным: Хелен кончила высшую школу и стала учительницей, тогда как Гледис не удалось кончить даже среднюю школу, и она работала в качестве мелкой служащей. Социальное окружение их отличалось во многих отношениях, хотя обе жили в городе. По росту, весу и т. д. у них разница невелика. Однако Хелен, жизнь которой сложилась удачнее, сохранилась физически лучше, чем ее близнячка. Различие в их умственном развитии очень заметно даже без применения тестов. Хелен, учительница, более слащава и любезна, более заботится о своей внешности и благоприятном впечатлении на окружающих. Гледис гораздо проще и мало озабочена производимым впечатлением.
Третья пара — Милфрид и Рут. Рано разлученные, они воспитывались в совсем разных семьях. Первая жила в доме банкира города средней величины, получила хорошее образование и вращалась среди интеллигентных людей, посещавших дом ее приемных родителей. Рут попала в семью малообразованных людей из рабочих. Угнетенное положение в доме отразилось на ее характере: заторможенная, пугливая, недоверчивая, молчаливая, она говорила в лепечущей манере, неудачно выражаясь. Милфрид, наоборот, была общительна, свободно и бойко говорила. И хотя обе получили высшее образование, Милфрид согласно оценке с помощью тестов, была более развита.
Эти 3 пары принадлежат к тем, которые из числа изучавшихся 20 пар имели ряд очень значительных различий в условиях жизни, в зависимости от чего можно поставить некоторые внутрипарные различия этих близнецов, особенно психики.
 Четвертая пара — мужчины 26 лет, Джемс и Рис (рис. 184). Дети углекопа, эти близнецы были рано разлучены и до 26 лет не виделись. Они воспитывались в разных семьях: один у родителей отца, другой у родителей рано умершей матери. Джемс попал в зажиточную деловую семью в маленький город. Впоследствии он стал инженером, женился и имел двух детей. Жизнь его не отличалась широким кругозором. Рис вырос в рабочей семье в горах Теннеси. Дед его не раз менял место работы. Рис не привык к регулярной работе, его влекла «вольная» жизнь гор, не раз сидел в тюрьме. Детей не имел. Он не закончил среднюю школу, и образование его было значительно ниже, чем его близнеца. Социальная среда, в которой каждый из них жил, тоже была заметно различная. Внешне эти близнецы были очень похожи, хотя и имели ряд мелких различий: Джемс весил примерно на 14 фунтов больше Риса, голова его была немного шире, чем голова Риса. Джемс был правша, Рис — левша. С помощью тестов был установлен значительно более высокий уровень образованности Джемса но сравнению с Рисом. Однако с помощью соответствующих тестов («Downey Will — Temperament Test»), характеризующих их «волю» и «темперамент», обнаружено их удивительное сходство (рис. 185). Оба проявили большую стойкость в противодействии, решительность суждения, сравнительно низкую «координацию импульсов» и «волевую настойчивость». Моторика обоих оказалась сравнительно медленной и слабой. Внешнее поведение их похоже: это очень спокойные, любезные люди.
Четвертая пара — мужчины 26 лет, Джемс и Рис (рис. 184). Дети углекопа, эти близнецы были рано разлучены и до 26 лет не виделись. Они воспитывались в разных семьях: один у родителей отца, другой у родителей рано умершей матери. Джемс попал в зажиточную деловую семью в маленький город. Впоследствии он стал инженером, женился и имел двух детей. Жизнь его не отличалась широким кругозором. Рис вырос в рабочей семье в горах Теннеси. Дед его не раз менял место работы. Рис не привык к регулярной работе, его влекла «вольная» жизнь гор, не раз сидел в тюрьме. Детей не имел. Он не закончил среднюю школу, и образование его было значительно ниже, чем его близнеца. Социальная среда, в которой каждый из них жил, тоже была заметно различная. Внешне эти близнецы были очень похожи, хотя и имели ряд мелких различий: Джемс весил примерно на 14 фунтов больше Риса, голова его была немного шире, чем голова Риса. Джемс был правша, Рис — левша. С помощью тестов был установлен значительно более высокий уровень образованности Джемса но сравнению с Рисом. Однако с помощью соответствующих тестов («Downey Will — Temperament Test»), характеризующих их «волю» и «темперамент», обнаружено их удивительное сходство (рис. 185). Оба проявили большую стойкость в противодействии, решительность суждения, сравнительно низкую «координацию импульсов» и «волевую настойчивость». Моторика обоих оказалась сравнительно медленной и слабой. Внешнее поведение их похоже: это очень спокойные, любезные люди.
 Перед нами очень интересный материал, который, однако, остался, при современном уровне знаний, недостаточно раскрытым и понятым научно; в частности, физиология высшей нервной деятельности таких пар не изучалась, что несомненно представляло бы большой интерес.
Перед нами очень интересный материал, который, однако, остался, при современном уровне знаний, недостаточно раскрытым и понятым научно; в частности, физиология высшей нервной деятельности таких пар не изучалась, что несомненно представляло бы большой интерес.
 Большинство других пар этой группы были внутрипарно более похожи, чем только что рассмотренные близнецы. Из их числа в смысле внутрипарного сходства выделяется одна пара, позже найденная, это Лоис и Луиз 1920 г. рождения (рис. 186), о которых нам надо сказать несколько слов. Близнецы эти были разлучены вскоре после рождения и редко виделись до 18 лет, когда поступили в один и тот же университет. Среднее образование получили в разных городах: Луиз — в небольшом городке, а Лоис — в большом городе и несколько лучшей школе. Экономический и культурный уровень тех двух семейств, в которых росли близнецы, был приблизительно одинаковый. Как по ряду физических, так и психических черт эта пара ОБ оказалась исключительно похожей, не только самой похожей среди пар, выросших врозь, но и среди большинства пар ОБ, выросших вместе. Очевидно, различия среды этих близнецов не были достаточно велики, чтобы вызвать в них сколько нибудь существенные внутрипарные различия физического или психического порядка (Gardner a. Newman, 1940а). Не входя в подробности статистического анализа трех групп близнецов данного исследования, рассмотрим лишь одну из итоговых таблиц его (табл. 47), где приводятся коэффициенты корреляции по некоторым физическим и психическим показателям. Последние определялись по нескольким системам психологических тестов, на основании некоторых из них определялся «умственный возраст» и «коэффициент интеллигентности» (I. Q.), показатели весьма спорные.
Большинство других пар этой группы были внутрипарно более похожи, чем только что рассмотренные близнецы. Из их числа в смысле внутрипарного сходства выделяется одна пара, позже найденная, это Лоис и Луиз 1920 г. рождения (рис. 186), о которых нам надо сказать несколько слов. Близнецы эти были разлучены вскоре после рождения и редко виделись до 18 лет, когда поступили в один и тот же университет. Среднее образование получили в разных городах: Луиз — в небольшом городке, а Лоис — в большом городе и несколько лучшей школе. Экономический и культурный уровень тех двух семейств, в которых росли близнецы, был приблизительно одинаковый. Как по ряду физических, так и психических черт эта пара ОБ оказалась исключительно похожей, не только самой похожей среди пар, выросших врозь, но и среди большинства пар ОБ, выросших вместе. Очевидно, различия среды этих близнецов не были достаточно велики, чтобы вызвать в них сколько нибудь существенные внутрипарные различия физического или психического порядка (Gardner a. Newman, 1940а). Не входя в подробности статистического анализа трех групп близнецов данного исследования, рассмотрим лишь одну из итоговых таблиц его (табл. 47), где приводятся коэффициенты корреляции по некоторым физическим и психическим показателям. Последние определялись по нескольким системам психологических тестов, на основании некоторых из них определялся «умственный возраст» и «коэффициент интеллигентности» (I. Q.), показатели весьма спорные.
 Эти коэффициенты корреляции показывают степень внутрипарного сходства близнецов данных трех групп. Корреляция роста и размеров головы обеих групп ОБ мало различается; корреляция веса разлученных ОБ ближе к коэффициенту РБ, иначе говоря, вес больше зависит от условий среды, чем рост и размеры головы. Заметны различия психических свойств трех групп близнецов; в некоторых случаях коэффициент корреляции разлученных ОБ даже меньше, чем коэффициент РБ. Не доверяя в деталях выводам, сделанным на основании приведенных в таблице тестов, все же, однако, можно считать, что соответствующие коэффициенты корреляции, хотя бы очень приблизительно, говорят о значительных различиях умственного развития трех данных групп близнецов. Эти различия между двумя группами ОБ, по-видимому, зависят в какой-то мере от условий постнатального развития, что подтверждается рассмотрением конкретных пар, как например Мери и Мебл. Если это так, то, очевидно, различие среды разно отозвалось на различных признаках: рост и размеры головы мало изменились, вес — больше, а психические свойства еще больше, и притом, по-видимому, разные свойства в разной степени. Однако последнее очень неопределенно освещается материалом этого исследования. Вопрос о зависимости разных свойств психики от различных условий жизни здесь не разрешается; делается лишь попытка подойти к его постановке. В общих же чертах результаты исследования Ньюмена с сотрудниками совпадают с выводами других работ, о которых речь была выше: они говорят, что разные свойства человеческого организма в разной степени подвержены изменениям под влиянием факторов среды. Само понятие среды авторы пытаются уточнить и углубить, с симпатией относясь к высказыванию Дженнингса (Jennings, 1937): то, что может наследственность, среда тоже может.
Свое десятилетнее исследование Ньюмен с сотрудниками начал с надеждой приблизиться к решению общей проблемы взаимоотношения наследственности и среды. В процессе работы он убедился в том, что общая проблема распадается на множество меньших проблем, что нет одного общего решения этой основной проблемы и даже скорого решения для ряда подчиненных ей вопросов. Однако Ньюмен с сотрудниками добыли ряд интересных фактов, по мере возможности изучили и обобщили их и этим помогли науке, — если не в решении основной проблемы, то хотя бы в подходе к правильной постановке ее, ибо всякий талантливо и добросовестно проделанный опыт поучителен.
Эти коэффициенты корреляции показывают степень внутрипарного сходства близнецов данных трех групп. Корреляция роста и размеров головы обеих групп ОБ мало различается; корреляция веса разлученных ОБ ближе к коэффициенту РБ, иначе говоря, вес больше зависит от условий среды, чем рост и размеры головы. Заметны различия психических свойств трех групп близнецов; в некоторых случаях коэффициент корреляции разлученных ОБ даже меньше, чем коэффициент РБ. Не доверяя в деталях выводам, сделанным на основании приведенных в таблице тестов, все же, однако, можно считать, что соответствующие коэффициенты корреляции, хотя бы очень приблизительно, говорят о значительных различиях умственного развития трех данных групп близнецов. Эти различия между двумя группами ОБ, по-видимому, зависят в какой-то мере от условий постнатального развития, что подтверждается рассмотрением конкретных пар, как например Мери и Мебл. Если это так, то, очевидно, различие среды разно отозвалось на различных признаках: рост и размеры головы мало изменились, вес — больше, а психические свойства еще больше, и притом, по-видимому, разные свойства в разной степени. Однако последнее очень неопределенно освещается материалом этого исследования. Вопрос о зависимости разных свойств психики от различных условий жизни здесь не разрешается; делается лишь попытка подойти к его постановке. В общих же чертах результаты исследования Ньюмена с сотрудниками совпадают с выводами других работ, о которых речь была выше: они говорят, что разные свойства человеческого организма в разной степени подвержены изменениям под влиянием факторов среды. Само понятие среды авторы пытаются уточнить и углубить, с симпатией относясь к высказыванию Дженнингса (Jennings, 1937): то, что может наследственность, среда тоже может.
Свое десятилетнее исследование Ньюмен с сотрудниками начал с надеждой приблизиться к решению общей проблемы взаимоотношения наследственности и среды. В процессе работы он убедился в том, что общая проблема распадается на множество меньших проблем, что нет одного общего решения этой основной проблемы и даже скорого решения для ряда подчиненных ей вопросов. Однако Ньюмен с сотрудниками добыли ряд интересных фактов, по мере возможности изучили и обобщили их и этим помогли науке, — если не в решении основной проблемы, то хотя бы в подходе к правильной постановке ее, ибо всякий талантливо и добросовестно проделанный опыт поучителен.
Повторное обследование близнецов через 25 лет
Интересные данные обобщены Фершюром в его последней книге 1954 г. Ему удалось повторно в 1950 г. обследовать тех близнецов, которых он изучал раньше, в 1924 г., всего 150 пар, из них 100 пар ОБ и 50 пар РБ. При втором обследовании это были люди среднего и старого возраста. Сравнивая внутрипарно физические и психические особенности близнецов, можно было видеть, в той или иной мере отчетливо, как различные условия и события жизни за 25 лет отразились на свойствах каждого субъекта. Фершюр брал одни и те же антропометрические показатели, полученные в 1924 и 1950 гг., и вычислял процентное уклонение, графически изображая полученные данные (рис. 187 и др.). Рассмотрим некоторые примеры.
 ОБ Е78, женщины 48 лет. До 27 лет жили в достатке в сельской местности. Первая из близнячек вышла замуж за садовника и имела детей. Ее жизнь протекала в большом труде и заботах. Вторая сделалась повивальной бабкой, замужем не была и детей не имела. Черты лица первой тоньше, она молчаливее и сдержанней, чем вторая, более шумная и болтливая. Физически, однако, эти близнецы мало отличаются друг от друга (рис. 187). Более заметна разница между близнецами на другом примере (Е83), тоже женщин с заметными различиями биографии. Интересно, что и через 25 лет сохранились различия между ними (рис. 188).
ОБ Е78, женщины 48 лет. До 27 лет жили в достатке в сельской местности. Первая из близнячек вышла замуж за садовника и имела детей. Ее жизнь протекала в большом труде и заботах. Вторая сделалась повивальной бабкой, замужем не была и детей не имела. Черты лица первой тоньше, она молчаливее и сдержанней, чем вторая, более шумная и болтливая. Физически, однако, эти близнецы мало отличаются друг от друга (рис. 187). Более заметна разница между близнецами на другом примере (Е83), тоже женщин с заметными различиями биографии. Интересно, что и через 25 лет сохранились различия между ними (рис. 188).
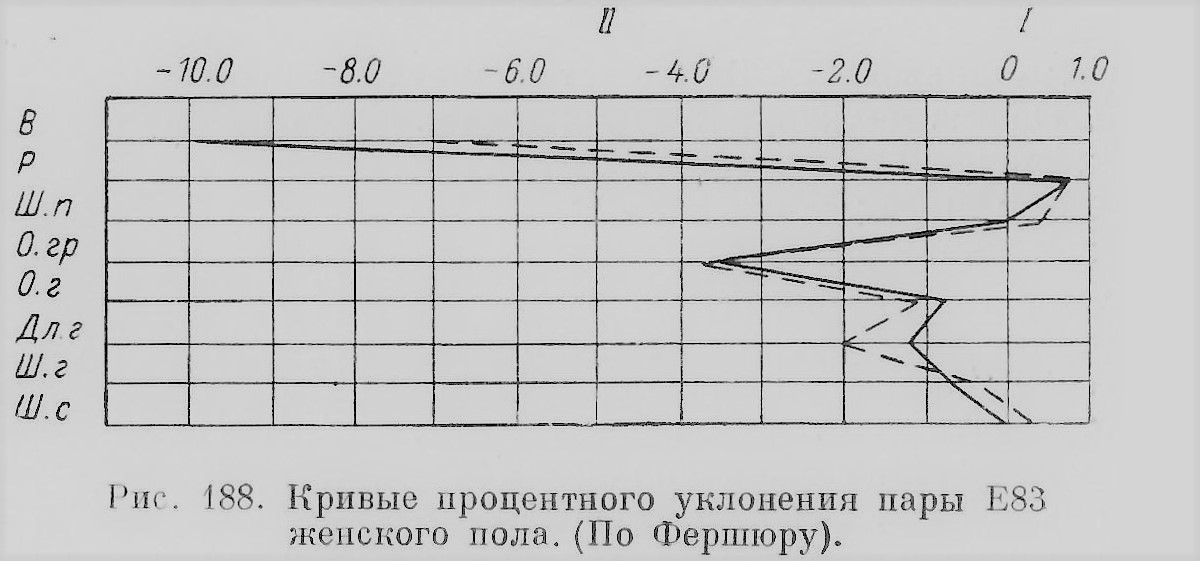 Встречаются случаи, когда близнецы с годами становятся более похожи, как например пара ОБ мужчин (Е84), один гравер, другой столяр, с относительно небольшими различиями в условиях жизни (рис. 189).
Встречаются случаи, когда близнецы с годами становятся более похожи, как например пара ОБ мужчин (Е84), один гравер, другой столяр, с относительно небольшими различиями в условиях жизни (рис. 189).
 Пример обратного представляют женщины-близнецы (Е108) 36 лет с заметно непохожим течением жизни. Обе из мелкобуржуазной среды, очень различно вышли замуж: первая — удачно, живет зажиточно и имеет троих детей; у второй муж пьяница, из-за которого она много страдает, живет беднее своей близнячки, имеет одного ребенка. Вторая долго болела, вследствие чего потеряла в весе и стала слабее первой, тогда как раньше была сильнее и даже выше ростом, чем первая (рис. 190).
Пример обратного представляют женщины-близнецы (Е108) 36 лет с заметно непохожим течением жизни. Обе из мелкобуржуазной среды, очень различно вышли замуж: первая — удачно, живет зажиточно и имеет троих детей; у второй муж пьяница, из-за которого она много страдает, живет беднее своей близнячки, имеет одного ребенка. Вторая долго болела, вследствие чего потеряла в весе и стала слабее первой, тогда как раньше была сильнее и даже выше ростом, чем первая (рис. 190).
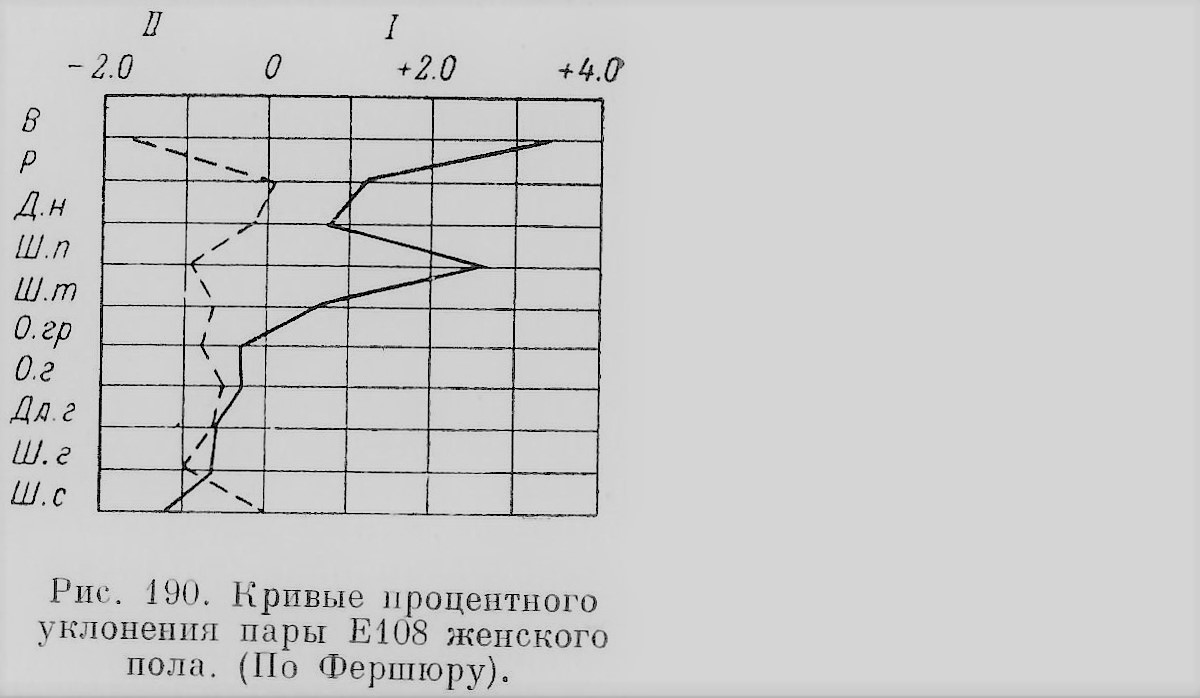 До известной степени аналогичный пример представляют ОБ мужского пола (Е64) в возрасте 42 лет. В детстве они были исключительно похожи, так что даже мать с трудом их различала. Оба посвятили себя архитектуре, но учились неодинаково. Второй окончил курс значительно раньше. Во время войны их судьба тоже была различна — второй был в плену и т. д. Жизнь первого в общем была труднее, он страдает астмой. Лицом они заметно различаются (рис. 191). Второй в общем несколько крупнее первого. Обмеры 1950 г. по ряду признаков отошли от данных 1924 г. (рис. 192).
До известной степени аналогичный пример представляют ОБ мужского пола (Е64) в возрасте 42 лет. В детстве они были исключительно похожи, так что даже мать с трудом их различала. Оба посвятили себя архитектуре, но учились неодинаково. Второй окончил курс значительно раньше. Во время войны их судьба тоже была различна — второй был в плену и т. д. Жизнь первого в общем была труднее, он страдает астмой. Лицом они заметно различаются (рис. 191). Второй в общем несколько крупнее первого. Обмеры 1950 г. по ряду признаков отошли от данных 1924 г. (рис. 192).
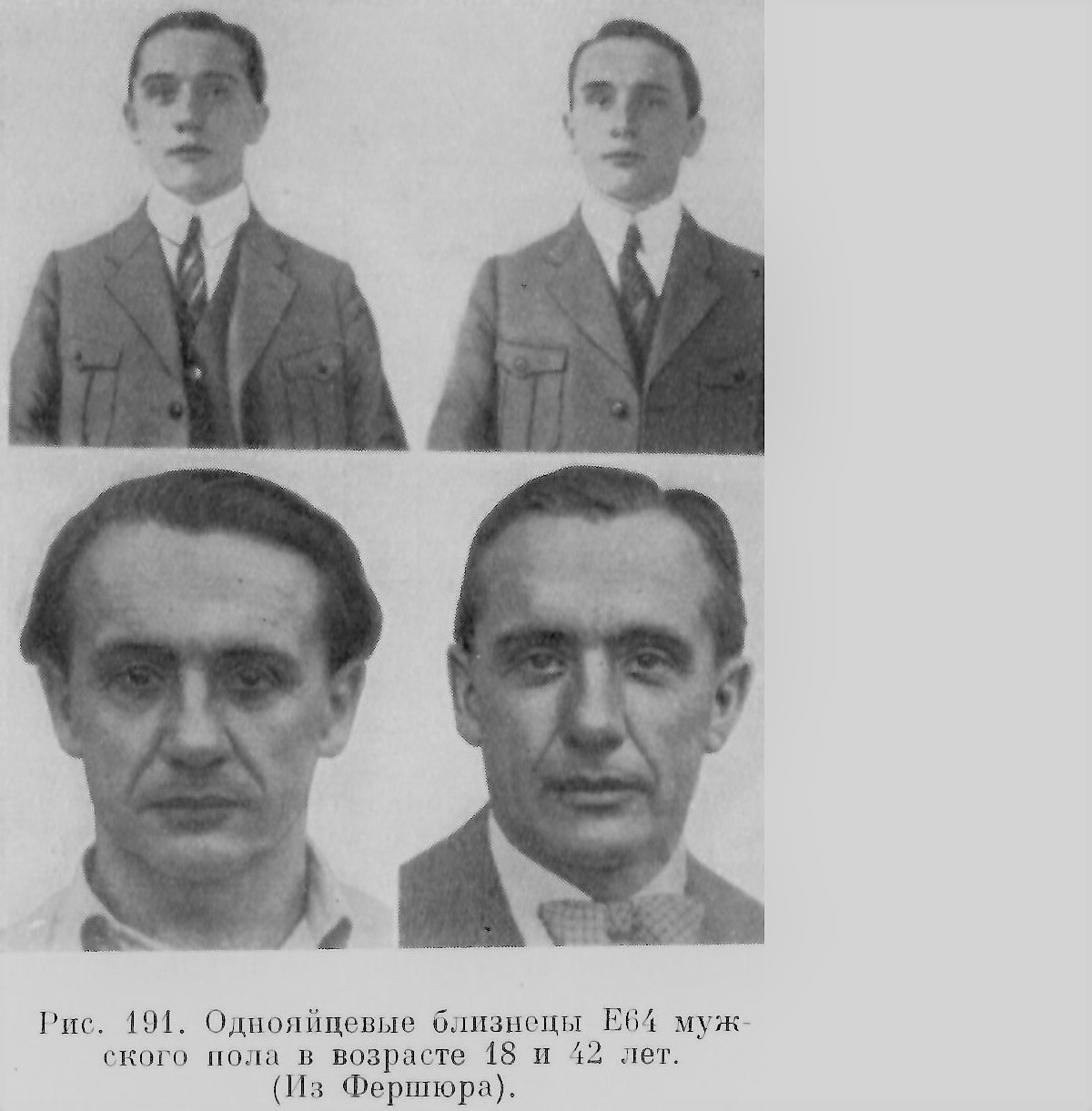
 Обобщая полученные результаты, Фершюр находит в новом материале подтверждение старого вывода о том, что различные признаки в разной мере изменчивы под влиянием среды. Так, вес очень изменчив, тогда как рост, наоборот, относительно мало изменчив. Череп более изменчив только в определенном возрасте и то в ограниченной степени и т. д.
Если взять средне процентное уклонение для 9 промеров тела и 5 промеров головы, отбросив вес тела, то можно констатировать, что в общем для всех ОБ разница с возрастом несколько увеличилась: в 1924 г. она была 0.80, а в 1950 — 0.93. Аналогичная разница для всех РБ оказывается несколько больше: 1.54 в 1924 г., но возрастает в 1950 г. только до 1.75, т. е. немногим больше, чем у ОБ.
Пытаясь глубже анализировать факторы жизни, могущие вызвать внутрипарные различия у близнецов, Фершюр выделяет их три для близнецов женского пола: 1) среду в обычном смысле слова, как-то: место обитания, социальные условия, занятие, питание и т. п.; 2) болезни; 3) рождение детей, так как последнее обстоятельство, по наблюдениям Фершюра, в очень большой степени влияет на судьбу женщины. Фершюр пытается группировать свой материал в зависимости от того, «похожи» или «непохожи» в общем эти три фактора для разных пар. Он отмечает, что если относительно болезнен и детей еще можно судить с известной степенью определенности, то относительно сходства среды судить значительно труднее. Его группировка, особенно в отношении последнего фактора, конечно, может вызвать сомнения и возражения, а потому и общие выводы, которые он делает на этой основе, далеко не достаточно убедительны. Мы потому и ограничиваемся одним лишь из его выводов, графически изображенным на рис. 193. Дискордантность по трем или двум из упомянутых факторов очень заметно сказывается в общем на весе, мало на объеме груди и совсем незначительно па росте, ширине плеч и других признаках.
Обобщая полученные результаты, Фершюр находит в новом материале подтверждение старого вывода о том, что различные признаки в разной мере изменчивы под влиянием среды. Так, вес очень изменчив, тогда как рост, наоборот, относительно мало изменчив. Череп более изменчив только в определенном возрасте и то в ограниченной степени и т. д.
Если взять средне процентное уклонение для 9 промеров тела и 5 промеров головы, отбросив вес тела, то можно констатировать, что в общем для всех ОБ разница с возрастом несколько увеличилась: в 1924 г. она была 0.80, а в 1950 — 0.93. Аналогичная разница для всех РБ оказывается несколько больше: 1.54 в 1924 г., но возрастает в 1950 г. только до 1.75, т. е. немногим больше, чем у ОБ.
Пытаясь глубже анализировать факторы жизни, могущие вызвать внутрипарные различия у близнецов, Фершюр выделяет их три для близнецов женского пола: 1) среду в обычном смысле слова, как-то: место обитания, социальные условия, занятие, питание и т. п.; 2) болезни; 3) рождение детей, так как последнее обстоятельство, по наблюдениям Фершюра, в очень большой степени влияет на судьбу женщины. Фершюр пытается группировать свой материал в зависимости от того, «похожи» или «непохожи» в общем эти три фактора для разных пар. Он отмечает, что если относительно болезнен и детей еще можно судить с известной степенью определенности, то относительно сходства среды судить значительно труднее. Его группировка, особенно в отношении последнего фактора, конечно, может вызвать сомнения и возражения, а потому и общие выводы, которые он делает на этой основе, далеко не достаточно убедительны. Мы потому и ограничиваемся одним лишь из его выводов, графически изображенным на рис. 193. Дискордантность по трем или двум из упомянутых факторов очень заметно сказывается в общем на весе, мало на объеме груди и совсем незначительно па росте, ширине плеч и других признаках.
 Очень интересны данные относительно болезней близнецов. Оказалось, что если учитывать только серьезные болезни, то среди ОБ обнаруживается подавляющее число дискордантных пар, т. е. таких, в которых один субъект болен, а другой здоров. Нечто подобное наблюдается и у РБ. Подробнее об этих данных речь была выше (см. главу двенадцатую «Болезни близнецов в онтогенезе»). Для общегенетических соображений эти данные Фершюра о болезнях близнецов очень важны. Он сам считает, что они исключают допущение какой-либо специальной наследственности этих болезней. Но, с другой стороны, он пытается показать, что они не зависят и от условий среды. Такая попытка совсем неубедительна; неясность вопроса о зависимости ряда болезней от условий среды, малая изученность еще не аннулируют вопроса, а наоборот, требуется его четкая постановка и углубленное исследование.
Интересные данные Фершюра о сроках смерти близнецов рассматривались выше.
Фершюр занимался и психикой близнецов. Он отверг «психометрию» с помощью тестов, которой раньше пользовался, и ограничился просто описанием отдельных случаев. Его общие выводы о существовании разных типов людей в смысле их «устойчивости» к влиянию среды как в физическом, так и психическом отношении, довольно неопределенны и мало разработаны. Его заключительный вывод о том, что факторами наследственности и среды не исчерпываются те «силы», которыми определяются жизнь и судьба человека, методологически неясен и недостаточно обоснован. Обширный и тщательно собранный материал, опубликованный в монографии Фершюра, может и должен рассматриваться не только с точки зрения автора, которая не всегда выдерживает критику. Новый материал в таком же роде собран учеником Фершюра, Дуисом (Duis, 1956). Опубликованная часть его касается различных болезней. Конечно, предпринятая Фершюром интересная работа не по силам одному исследователю, даже имеющему помощников. Изучаемая группа близнецов должна обследоваться чаще, чем 2 раза за 25 лет, в ряде моментов должна исследоваться глубже и разностороннее, чем это делалось до сих пор, с привлечением экспериментов, изучаться должны также родственники близнецов и т. д.
Очень интересны данные относительно болезней близнецов. Оказалось, что если учитывать только серьезные болезни, то среди ОБ обнаруживается подавляющее число дискордантных пар, т. е. таких, в которых один субъект болен, а другой здоров. Нечто подобное наблюдается и у РБ. Подробнее об этих данных речь была выше (см. главу двенадцатую «Болезни близнецов в онтогенезе»). Для общегенетических соображений эти данные Фершюра о болезнях близнецов очень важны. Он сам считает, что они исключают допущение какой-либо специальной наследственности этих болезней. Но, с другой стороны, он пытается показать, что они не зависят и от условий среды. Такая попытка совсем неубедительна; неясность вопроса о зависимости ряда болезней от условий среды, малая изученность еще не аннулируют вопроса, а наоборот, требуется его четкая постановка и углубленное исследование.
Интересные данные Фершюра о сроках смерти близнецов рассматривались выше.
Фершюр занимался и психикой близнецов. Он отверг «психометрию» с помощью тестов, которой раньше пользовался, и ограничился просто описанием отдельных случаев. Его общие выводы о существовании разных типов людей в смысле их «устойчивости» к влиянию среды как в физическом, так и психическом отношении, довольно неопределенны и мало разработаны. Его заключительный вывод о том, что факторами наследственности и среды не исчерпываются те «силы», которыми определяются жизнь и судьба человека, методологически неясен и недостаточно обоснован. Обширный и тщательно собранный материал, опубликованный в монографии Фершюра, может и должен рассматриваться не только с точки зрения автора, которая не всегда выдерживает критику. Новый материал в таком же роде собран учеником Фершюра, Дуисом (Duis, 1956). Опубликованная часть его касается различных болезней. Конечно, предпринятая Фершюром интересная работа не по силам одному исследователю, даже имеющему помощников. Изучаемая группа близнецов должна обследоваться чаще, чем 2 раза за 25 лет, в ряде моментов должна исследоваться глубже и разностороннее, чем это делалось до сих пор, с привлечением экспериментов, изучаться должны также родственники близнецов и т. д.
Глава четырнадцатая МЕТОДЫ ОПРЕДЕЛЕНИЯ ТИПА БЛИЗНЕЦОВ
На каком основании можно решить, что данная пара близнецов принадлежит к одному из двух типов — ОБ или РБ? Это основной вопрос для исследовательской работы с близнецами. Ни Гальтон (Galton, 1875), ни Торндайк (Thorndike, 1905) не могли на него ответить отчетливо, так как этот вопрос не был поставлен достаточно ясно даже в начале нашего века. Только в 20-е годы (Apert, 1923; Siemens, 1924) он был выяснен настолько, что после этого открылись широкие перспективы изучения близнецов и появилось множество исследований их. Основным объектом был человек. Принадлежность близнецов к тому или другому типу или, как иногда для краткости говорят, диагноз типа близнецов, решалась раньше на основании их зародышевых оболочек. Было принято считать, что ОБ имеют один хорион, а РБ — разные хорионы. Мы знаем, что этот критерий благодаря новым исследованиям был заново рассмотрен и значительно ограничен, так как установлены случаи двухориальных ОБ и т. д. (см. главу четвертую). Однако этим значение оболочек как критерия диагноза типа близнецов не вовсе отвергается, а лишь ограничивается — ему более не придается безусловно решающего значения, а только подсобное, наряду с другими критериями, о которых речь будет ниже. Определение типа близнецов с помощью оболочек не всегда возможно, даже редко когда возможно, так как чаще приходится иметь дело с близнецами, особенно взрослыми, относительно оболочек которых или вовсе нет сведений, или имеются сведения неточные и недостаточно достоверные. Поэтому для решения вопроса о типе пары близнецов, об оболочках которых ничего достоверно не известно, надо иметь другой метод, от оболочек не зависящий. Таковым и является метод, называемый «полисимптоматическим методом сходства» или просто «методом сходства». Он состоит в сравнении близнецов по ряду признаков и выяснении внутрипарного сходства или различия по ним. С помощью этого метода можно установить тип близнецов почти всегда достоверно, как показали многочисленные проверки его (Verschuer, 1932; Ардашников и др., 1936а, 1936б; Lotze, 1937; Newman, 1940b; Gedda, 1951, и др.). Какие признаки надо брать для определения типа близнецов? Этот вопрос решается в связи с тем основным различием, которое имеется между двумя типами близнецов, ОБ и РБ. ОБ по своей природе должны иметь сходство генотипа, исключительно большое наследственное сходство, максимальное между двумя людьми. РБ, как братья и сестры, должны иметь меньшее сходство наследственности, чем ОБ. Это различие между обоими типами близнецов должно проявляться и проявляется в том, что по множеству признаков ОБ внутрипарно более похожи, чем РБ. Вышеизложенный материал достаточно красноречиво говорит об этом как о несомненном факте. Для диагноза типа близнецов, очевидно, надо брать не все возможные для исследования свойства и признаки, а лишь те, которые отчетливее других могут показать наследственное сходство пары ОБ. Это должны быть такие признаки, которые меньше других подвергнуты изменчивости под влиянием факторов среды. Мы уже с такими признаками знакомы. К ним относятся: серологические свойства крови (группы крови, факторы М и N, Rh и др.), пигментация глаз, кожи и волос, форма волос и отчасти распределение волосяного покрова, кожный рельеф (отпечатки пальцев, ладоней и ступней), некоторые морфологические особенности (форма носа, губ, ушей, кистей рук и т. п.), форма, цвет и расположение зубов и др.. Проверка пригодности ряда этих признаков для диагностики типа близнецов дала положительные результаты. В качестве примера рассмотрим таблицу (табл. 48), показывающую распределение пар близнецов по числу дискордантных признаков (Ардашников и др., 1936). Мы видим, что среди ОБ подавляющее большинство пар (90) оказывается конкордантным, т. е. не имеет различии по всем взятым для диагноза признакам. 34 пары имеют различия, т. е. дискордантны, по одному признаку, 11 — по двум и только 2 пары — по трем признакам. Наоборот, РБ, как минимум, имеют различие по трем признакам, а наибольшее число пар РБ имеют различия по восьми признакам. Из табл. 48 хорошо видно своеобразие обоих типов близнецов, и вместе с тем становится ясным, что действительно существует только два типа близнецов, нет третьего, промежуточного, и нет промежуточных форм близнецов, как думали прежде некоторые авторы (Ардашников и др., 1936б).
Аналогичную картину получил Ньюмен с сотрудниками (Newman et al., 1937); она отчетливо видна на ряде кривых, изображающих внутрипарную разность обоих типов близнецов по отдельным признакам, как рост, вес и т. д. (см. рис. 133).
В табл. 48 выделено 7 «сомнительных» пар. Это пары, тип которых с трудом поддается выяснению, так как у этих близнецов имеются некоторые заметные различия по признакам, которые обычно одинаковы у ОБ, например по цвету глаз, форме бровей и т. д. — различия, которые редко, но встречаются между правой и левой половиной одного и того же индивидуума. В других случаях различия вызваны болезнью, перенесенной только одним из партнеров. В большинстве случаев эти «сомнительные» пары относятся к типу ОБ (Siemens, 1927; Ардашников и др., 1936, и др.). Делались попытки математически выразить вероятность принадлежности данной пары близнецов к одному из двух типов на основании сходства по определенному числу отобранных для диагноза признаков (Бунак, 1926; Rife, 1933b; Essen-Moller, 1941, и др.). Но для опытного диагноста близнецов эти формулы мало что дают, и можно вполне обходиться без них.
Различные исследователи определяли, как часто среди ОБ и РБ встречаются различия по употребляемым для диагноза признакам. В качестве примеров приведем таблицу, составленную советскими учеными (Ардашников и др., 1936) для признаков, которыми они пользовались для диагноза, где показан процент близнецов с внутрипарными различиями к общему числу близнецов изученной группы (табл. 49).
Мы видим, что среди ОБ подавляющее большинство пар (90) оказывается конкордантным, т. е. не имеет различии по всем взятым для диагноза признакам. 34 пары имеют различия, т. е. дискордантны, по одному признаку, 11 — по двум и только 2 пары — по трем признакам. Наоборот, РБ, как минимум, имеют различие по трем признакам, а наибольшее число пар РБ имеют различия по восьми признакам. Из табл. 48 хорошо видно своеобразие обоих типов близнецов, и вместе с тем становится ясным, что действительно существует только два типа близнецов, нет третьего, промежуточного, и нет промежуточных форм близнецов, как думали прежде некоторые авторы (Ардашников и др., 1936б).
Аналогичную картину получил Ньюмен с сотрудниками (Newman et al., 1937); она отчетливо видна на ряде кривых, изображающих внутрипарную разность обоих типов близнецов по отдельным признакам, как рост, вес и т. д. (см. рис. 133).
В табл. 48 выделено 7 «сомнительных» пар. Это пары, тип которых с трудом поддается выяснению, так как у этих близнецов имеются некоторые заметные различия по признакам, которые обычно одинаковы у ОБ, например по цвету глаз, форме бровей и т. д. — различия, которые редко, но встречаются между правой и левой половиной одного и того же индивидуума. В других случаях различия вызваны болезнью, перенесенной только одним из партнеров. В большинстве случаев эти «сомнительные» пары относятся к типу ОБ (Siemens, 1927; Ардашников и др., 1936, и др.). Делались попытки математически выразить вероятность принадлежности данной пары близнецов к одному из двух типов на основании сходства по определенному числу отобранных для диагноза признаков (Бунак, 1926; Rife, 1933b; Essen-Moller, 1941, и др.). Но для опытного диагноста близнецов эти формулы мало что дают, и можно вполне обходиться без них.
Различные исследователи определяли, как часто среди ОБ и РБ встречаются различия по употребляемым для диагноза признакам. В качестве примеров приведем таблицу, составленную советскими учеными (Ардашников и др., 1936) для признаков, которыми они пользовались для диагноза, где показан процент близнецов с внутрипарными различиями к общему числу близнецов изученной группы (табл. 49).
 Для сравнения приведем еще таблицу признаков, которыми пользуется Фершюр (табл. 50, — Lotze, 1937).
Для сравнения приведем еще таблицу признаков, которыми пользуется Фершюр (табл. 50, — Lotze, 1937).
 Мы видим, что в списке немецких ученых фигурируют несколько иные признаки, чем у советских ученых; по признакам же, совпадающим у тех и других, процент дискордантных случаев иной, например, по цвету глаз у советских ученых имеется 4.6% дискордантных пар, а у немецких — 0%, но зато 13% с небольшими вариациями, и т. д. Это зависит, конечно, от материала и приемов оценки сходства-различия. Но главное заключается в том, что по степени дискордантности по всем признакам обеих таблиц РБ резко превосходят ОБ.
Некоторые исследователи добавляют еще иные признаки и заменяют часть из приведенных в этих таблицах признаков другими. Так, Ньюмен, например, придает большое значение морфологии лица и кожным узорам пальцев и ладоней как диагностическим признакам, другие — цвету и форме зубов, психическим особенностям, некоторым патологическим признакам и аномалиям, там, где они выражены и известны как наследственные (аномалии прикуса, глаз, шестипалость и т. д.). Некоторые ученые ограничиваются сравнительно немногими признаками, которые считают наиболее важными, например следующими: серологическими показателями крови, отпечатками кожных узоров, цветом и рефракцией глаз, формой и цветом волос, завитком волос на темени, цветом кожи, ресницами и бровями (например, Мак-Артур при диагнозе пятерни Дионн).
В общем же большинство исследователей разных стран, работающих с близнецами, в основном пользуется одними и теми же признаками для диагноза типа, достаточно уже проверенными опытом мировой науки на тысячах пар близнецов.
Из природы этих признаков явствует, что в массе они не применимы к детям до второй половины первого года жизни, так как у младенцев в первые месяцы жизни трудно судить о цвете глаз, кожи и иногда даже волос, о бровях и ресницах, трудно делать отпечатки пальцев и т. д., а вес, рост, форма головы и т. п. признаки еще носят явные последствия условий утробной жизни. Здесь обнаруживается неизбежное бессилие «метода сходства», потому что большинство признаков, которыми пользуется этот метод, в таком раннем возрасте еще недоступно изучению. Пока что выделено было только три признака, которыми, по-видимому, можно с успехом пользоваться у новорожденных — это цвет волос, группы крови и форма кожных капилляров (Rohlfs, 1928).
Далее надо отметить, что для некоторых рас известные признаки из приведенных оказываются неприменимыми. Так, например, вся серия признаков окраски кожи, глаз, волос, а также форма последних оказывается непригодной для китайцев или негров: РБ, как и ОБ, по ним одинаково конкордантны. Для этих рас особенно важное значение приобретают кожные узоры как диагностический признак. Вышеприведенные ряды признаков выработаны на европейских расах и применимы прежде всего к ним.
Диагноз типа ставится следующим образом: если пара близнецов одинакового пола (разумеется, если пол разный, то это без дальнейших наблюдений РБ) похожа до тождества или почти до тождества по ряду наиболее употребительных признаков, то эта пара ОБ. Если по одному-двум признакам близнецы внутрипарно безусловно различны, дискордантны, например, по группам крови, цвету глаз и волос и т. д., то они РБ. Разумеется, что небольшое различие оттенка цвета волос и формы носа не является доказательством разнояйцевости, если целый ряд других признаков свидетельствует в пользу однояйцевости. Встречаются, правда довольно редко, сомнительные случаи, которые даже опытные специалисты затрудняются отнести к тому или другому типу, как например пара девочек, изображенная на рис. 194, которых такой знаток близнецов, как Сименс (Siemens, 1927), не мог диагносцировать.
Мы видим, что в списке немецких ученых фигурируют несколько иные признаки, чем у советских ученых; по признакам же, совпадающим у тех и других, процент дискордантных случаев иной, например, по цвету глаз у советских ученых имеется 4.6% дискордантных пар, а у немецких — 0%, но зато 13% с небольшими вариациями, и т. д. Это зависит, конечно, от материала и приемов оценки сходства-различия. Но главное заключается в том, что по степени дискордантности по всем признакам обеих таблиц РБ резко превосходят ОБ.
Некоторые исследователи добавляют еще иные признаки и заменяют часть из приведенных в этих таблицах признаков другими. Так, Ньюмен, например, придает большое значение морфологии лица и кожным узорам пальцев и ладоней как диагностическим признакам, другие — цвету и форме зубов, психическим особенностям, некоторым патологическим признакам и аномалиям, там, где они выражены и известны как наследственные (аномалии прикуса, глаз, шестипалость и т. д.). Некоторые ученые ограничиваются сравнительно немногими признаками, которые считают наиболее важными, например следующими: серологическими показателями крови, отпечатками кожных узоров, цветом и рефракцией глаз, формой и цветом волос, завитком волос на темени, цветом кожи, ресницами и бровями (например, Мак-Артур при диагнозе пятерни Дионн).
В общем же большинство исследователей разных стран, работающих с близнецами, в основном пользуется одними и теми же признаками для диагноза типа, достаточно уже проверенными опытом мировой науки на тысячах пар близнецов.
Из природы этих признаков явствует, что в массе они не применимы к детям до второй половины первого года жизни, так как у младенцев в первые месяцы жизни трудно судить о цвете глаз, кожи и иногда даже волос, о бровях и ресницах, трудно делать отпечатки пальцев и т. д., а вес, рост, форма головы и т. п. признаки еще носят явные последствия условий утробной жизни. Здесь обнаруживается неизбежное бессилие «метода сходства», потому что большинство признаков, которыми пользуется этот метод, в таком раннем возрасте еще недоступно изучению. Пока что выделено было только три признака, которыми, по-видимому, можно с успехом пользоваться у новорожденных — это цвет волос, группы крови и форма кожных капилляров (Rohlfs, 1928).
Далее надо отметить, что для некоторых рас известные признаки из приведенных оказываются неприменимыми. Так, например, вся серия признаков окраски кожи, глаз, волос, а также форма последних оказывается непригодной для китайцев или негров: РБ, как и ОБ, по ним одинаково конкордантны. Для этих рас особенно важное значение приобретают кожные узоры как диагностический признак. Вышеприведенные ряды признаков выработаны на европейских расах и применимы прежде всего к ним.
Диагноз типа ставится следующим образом: если пара близнецов одинакового пола (разумеется, если пол разный, то это без дальнейших наблюдений РБ) похожа до тождества или почти до тождества по ряду наиболее употребительных признаков, то эта пара ОБ. Если по одному-двум признакам близнецы внутрипарно безусловно различны, дискордантны, например, по группам крови, цвету глаз и волос и т. д., то они РБ. Разумеется, что небольшое различие оттенка цвета волос и формы носа не является доказательством разнояйцевости, если целый ряд других признаков свидетельствует в пользу однояйцевости. Встречаются, правда довольно редко, сомнительные случаи, которые даже опытные специалисты затрудняются отнести к тому или другому типу, как например пара девочек, изображенная на рис. 194, которых такой знаток близнецов, как Сименс (Siemens, 1927), не мог диагносцировать.
 Подобные пары или РБ, имеющие ряд одинаковых признаков в силу сходства их наследственности по ним, что может зависеть от сходства родителей, например по цвету волос, глаз и т. д., или же это ОБ, у которых в силу особых обстоятельств, например утробной жизни, одни из партнеров получил заметные от другого отличия. В таких случаях, если это действительно ОБ, конечно, важно увидеть типически общее данной пары близнецов, не взирая на частные, но нерешающие, различия, как например небольшие различия морфологии или окраски (см. Бунак, 1926).
Ставился вопрос: какие шансы имеет пара РБ быть столь же похожей, как пара ОБ? Иначе говоря, насколько вероятна такая степень внутрипарного сходства РБ, что их можно принять за ОБ. Райф (Rife, 1933а) специально занимался этим вопросом, пользуясь всего только восемью признаками (группы крови, факторы М и N, наличие-отсутствие волос на третьей фаланге пальцев, вкусовая реакция на РТС, окраска радужной оболочки, число дактилоскопических полосок, I. Q., рост). Для каждого из этих признаков он вычислил вероятность их сходства у пары РБ, имея чисто гипотетические основы для этих расчетов. Для разных случаев он нашел шансы того, что данная пара не ОБ, а РБ, такого порядка: 1 на 480 ООО, 1 на 319 999 и т. д. Как бы приблизительны не были эти расчеты, все же они отражают действительность в том смысле, что вероятность для пары РБ быть столь похожей, что ее можно принять за ОБ, крайне мала, настолько, что вряд ли может реально встретиться такой случай. Если учесть не восемь признаков, а большее число их, так как в действительности их, конечно, не так мало, то шансы на сходство всех этих признаков у пары РБ становятся еще меньше. Это говорит в пользу бесспорной практической пригодности «метода сходства» для диагноза типов близнецов, и всякий, кто с этим вопросом имел дело, не сомневается в его научной ценности.
«Метод сходства», позволяющий достаточно убедительно различать оба типа близнецов, является основой близнецового метода генетики.
Подобные пары или РБ, имеющие ряд одинаковых признаков в силу сходства их наследственности по ним, что может зависеть от сходства родителей, например по цвету волос, глаз и т. д., или же это ОБ, у которых в силу особых обстоятельств, например утробной жизни, одни из партнеров получил заметные от другого отличия. В таких случаях, если это действительно ОБ, конечно, важно увидеть типически общее данной пары близнецов, не взирая на частные, но нерешающие, различия, как например небольшие различия морфологии или окраски (см. Бунак, 1926).
Ставился вопрос: какие шансы имеет пара РБ быть столь же похожей, как пара ОБ? Иначе говоря, насколько вероятна такая степень внутрипарного сходства РБ, что их можно принять за ОБ. Райф (Rife, 1933а) специально занимался этим вопросом, пользуясь всего только восемью признаками (группы крови, факторы М и N, наличие-отсутствие волос на третьей фаланге пальцев, вкусовая реакция на РТС, окраска радужной оболочки, число дактилоскопических полосок, I. Q., рост). Для каждого из этих признаков он вычислил вероятность их сходства у пары РБ, имея чисто гипотетические основы для этих расчетов. Для разных случаев он нашел шансы того, что данная пара не ОБ, а РБ, такого порядка: 1 на 480 ООО, 1 на 319 999 и т. д. Как бы приблизительны не были эти расчеты, все же они отражают действительность в том смысле, что вероятность для пары РБ быть столь похожей, что ее можно принять за ОБ, крайне мала, настолько, что вряд ли может реально встретиться такой случай. Если учесть не восемь признаков, а большее число их, так как в действительности их, конечно, не так мало, то шансы на сходство всех этих признаков у пары РБ становятся еще меньше. Это говорит в пользу бесспорной практической пригодности «метода сходства» для диагноза типов близнецов, и всякий, кто с этим вопросом имел дело, не сомневается в его научной ценности.
«Метод сходства», позволяющий достаточно убедительно различать оба типа близнецов, является основой близнецового метода генетики.
Глава пятнадцатая БЛИЗНЕЦОВЫЙ МЕТОД ГЕНЕТИКИ
Как уже выше говорилось, Гальтон (Galton, 1875) впервые высказал идею современного близнецового метода и пытался использовать близнецов для решения вопроса о роли факторов наследственности и среды для различных свойств человеческого организма. Как известно, Гальтон, убежденный в ведущей роли наследственности, «фатальности» ее, нашел и в близнецовом материале подтверждение этой мысли. Эту предвзятую установку вместе с близнецовым методом заимствовали в той или иной мере многие позднейшие исследователи нашего времени (Сименс, Фершюр, Дальберг и др.). Как мы увидим в дальнейшем, такая установка вовсе не обязательна для этого метода. Сущность близнецового метода генетики основывается па генотипическом сходстве пары ОБ и вытекающих отсюда возможностях различных внутрипарных и межпарных сравнений разных групп близнецов (Lotze, 1937; Newman, 1940с; Smith, 1949; Gedda, 1951; Rife, 1952a; Neel a. Schull, 1954, и др.). Близнецовый метод допускает три постулата. Во-первых, что пара однояйцевых близнецов имеет тождественный генотип, т. е. одинакова в отношении наследственности, тогда как пара разнояйцевых близнецов имеет неодинаковый генотип, конечно, не по всем признакам, что иногда забывается; иначе говоря, по некоторым свойствам и пара РБ может иметь одинаковую наследственность. Во-вторых, что среда, от воздействий которой могут возникать внутрипарные различия ОБ, может быть для одной какой-нибудь пары близнецов «одинаковой», а для другой пары — «разной», т. е. могут быть пары с внутрипарной «одинаковой» и внутрипарной «разной» средой. Наконец, третий постулат предполагает, что генезис всех структурных и функциональных свойств организмов зависит от взаимодействия двух родов факторов — факторов генотипа (наследственности) и факторов среды и что еще каких-либо иных факторов, влияющих на свойства организмов, не существует. Из этих допущений вытекает возможность существования разных групп близнецов: во-первых, ОБ с «одинаковой» средой для каждой пары, во-вторых, ОБ с «разной» средой для каждой пары, и, наконец, в-третьих, РБ с одинаковой средой для каждой пары. РБ с разной средой внутрипарно имеют меньшую методическую ценность. Эти категории близнецов можно сравнивать следующим образом: ОБ с одинаковой средой сравнивать с ОБ с разной средой — это дает возможность судить о влиянии среды; далее, ОБ с одинаковой средой сравнивать с РБ с одинаковой средой— эго дает возможность судить о роли наследственности. Такие сравнения делаются по возможности на большом материале, пригодном для статистической обработки. Так, если какой-нибудь признак, например вес, у ОБ с внутрипарно одинаковой средой проявляется конкордантно, а у ОБ с разной средой — дискордантно, то считается, что этот признак сравнительно легко меняется под влиянием среды, легче, напр., чем рост. Если же сравнивать серию ОБ, имеющую одинаковую среду, с серией РБ, тоже имеющей одинаковую среду, то большая дискордантность РБ будет зависеть от внутрипарной разницы генотипов, например дискордантность по цвету волос или числу дельт на отпечатках пальцев и т. д. Но возможны и другие способы сравнения, например внутрипарное сравнение одной или немногих пар ОБ в процессе их развития, заболевания и т. д. или в условиях эксперимента с одним из ОБ — метод «взаимоконтроля» Гизелла (см. главу одиннадцатую). При таком сравнении, полагая, что пара ОБ генетически тождественна, всякие различия считаются вызванными не генотипическими факторами, а факторами среды, например различия формы головы, веса, условных рефлексов и т. д. При сравнении отдельных пар ОБ, подвергнутых одинаковому воздействию среды, может быть установлено различие «пластичности» их генотипа (Rife, 1952а). Для подобных исследований полезно, следовательно, сравнение разных пар ОБ при параллельном внутрипарном их сравнении. Мы видели, что можно также сравнивать пары ОБ не только между собой и с парами РБ, но также с родственниками-неблизнецами разной степени близости родства (метод близнецовой семьи) и т. д. Как уже упоминалось, одним из основных обобщении, полученных путем изучения близнецов, был тот факт, что разные свойства животных и человека в разной степени изменчивы под влиянием воздействий среды. Это вызвало желание у ряда исследователей математическим путем установить «долю» наследственности и «долю» среды в определении какого-нибудь признака (Lenz-Verschuer, 1928; Игнатьев, 1936; Lotze, 1937; Newman et al., 1937; Dahlberg, 1942; Neel a. Schull, 1954, и др.). В качестве примера такого рода расчетов возьмем формулу Хольцингера:H = (CM — CD) / (100 — CD).Здесь Н обозначает долю наследственности, СМ — процент конкордантных пар ОБ, a CD — конкордантных пар РБ. Как известно, конкордантность РБ меньше, чем ОБ, что зависит от большего наследственного сходства ОБ. Поэтому приведенная разность и служит мерилом «доли» наследственности. Так вычисленное Н будет для роста 81%, для веса 78%, и т. д. (Newman, 1940с). Несмотря на то, что такие расчеты производят впечатление большой точности, они имеют спорную и весьма относительную ценность. Они построены прежде всего на упрощенно-механическом представлении о взаимодействии факторов среды и наследственности, по аналогии со сплавом двух металлов, долю каждого из которых можно в этом сплаве вычислить. Такое представление годится только в качестве метафоры, но не в качестве основы для количественных расчетов взаимоотношений наследственности и среды, которые гораздо сложнее и должны мыслиться диалектически в онтогенетическом аспекте. Далее, подобные расчеты обязательно должны были бы строиться на сериях ОБ и РБ без отбора (см. главу десятую), иначе искажается их отношение, соответствующее таковому в популяции, а в связи с этим получаются неверные цифры «доли» наследственности, что не раз имело место в литературе (ср. Neel a. Schull, 1954, и др.). Наконец, эти вычисления опираются на приведенные выше постулаты иногда без должной критики их, что также может опорочить их результаты. Поэтому мы не будем здесь углубляться в рассмотрение других формул и приемов подобных вычислений (Verschuer, 1927, 1932; Dahlberg, 1942; Rife, 1952a; Neel a. Schull, 1954, и др.). По-видимому, вопрос о роли наследственности в проявлении какого-нибудь признака у близнецов, если этот вопрос вообще может быть обработан математическим путем, требует пересмотра всех основ такого рода расчетов с новых позиций, пока еще отчетливо не сформулированных. Теперь обратимся к критическому рассмотрению постулатов близнецового метода. Первый из них утверждает тождество генотипа пары ОБ. Под генотипом в данном случае условимся понимать всю совокупность наследственной природы данного организма, независимо от того, где локализуются факторы наследственности, что они собой представляют и т. д. Некоторые авторы, различно аргументируя свои взгляды, выступали против этого тезиса близнецового метода (Dahlberg, 1926, 1943/44; Волоцкой, 1937; Bouterwek, 1943, и др.). Утверждение одних, что асимметрия и зеркальность пары ОБ зависит от различия их генотипов, остается недоказанным (см. главу седьмую). Утверждение других, что постулат тождества генотипов не доказан, конечно, справедливо, но и для обратного утверждения пока нет достаточных оснований. Разумеется, можно предположить, что какие-то элементы генотипа меняются то ли под влиянием условий среды, то ли «спонтанно», т. е. по каким-то пока неизвестным причинам, например возникают мутации или иные генотипические изменения у одного из ОБ, тогда как у другого они отсутствуют или оказываются иными. Однако конкретные изменения такого рода пока неизвестны, и вопрос этот остается открытым. Только будущие исследования могут принести ответ на него. Даже если будут доказаны какие-нибудь изменения генотипа в онтогенезе, происходящие у одного из ОБ и отсутствующие у другого, то возникнет вопрос, насколько такого рода изменения окажутся существенными и повлияют на разные категории признаков. Предположим, что будет, например, установлен случай соматической мутации у одного из близнецов, выражающийся в изменении окраски волос. Спрашивается, повлияет ли это изменение генотипа на свойства крови, на рост, рельеф кожи, цвет зубов, рефракцию хрусталика и т. д.? Едва ли. В таком случае, даже признав различие генотипа по цвету волос, можно спокойно работать с этой парой ОБ по рефракции хрусталика и другим признакам, имеющим в основе сходство генотипа этих близнецов. Вероятно, надо допустить, как исключительно редкую возможность, такое изменение генотипа одного из близнецов, которое затронет ряд признаков, изменит как-то весь генотип. Потому вряд ли предположением каких-либо изменений генотипа одного из пары ОБ можно опорочить первый постулат близнецового метода в целом. Наследственному сходству ОБ противопоставляется наследственное различие РБ. Не надо забывать, что оно далеко не всегда касается всех признаков. В ряде случаев РБ могут быть внутрипарно неследственно похожи, например по цвету волос, группе крови и т. д. Наличие огромного внутрипарного сходства ОБ по множеству самых различных морфологических и физиологических особенностей свидетельствует несомненно если не о тождестве их генотипа, то во всяком случае об исключительном сходстве генотипов обоих членов пары ОБ, большем, чем сходство между генотипами родителей и детей или братьев-сестер. До сих пор шла речь о внутрипарном сходстве фенотипа ОБ в силу генотипического сходства таких близнецов. На это можно возразить, сказав, что фенотипическое сходство может быть вызвано также факторами среды, одинаково действующими на обоих ОБ. В какой мере это так, зависит от того, о каких признаках идет речь. Влияние факторов среды можно проверить по крайней мере двояким способом: испытуемый признак изучить у пары ОБ, поставленной в разные условия среды, т. е. подвергнуть каждого из близнецов действию разных факторов среды, или же исследовать испытуемый признак на парах РБ, поставленных каждая в одинаковые условия среды, влияющие на изучаемый признак. Так можно подойти к решению вопроса о том, насколько сходство ОБ по данному признаку обусловлено сходством или различием факторов среды. Мы уже знаем, что ряд признаков в очень большой мере зависит от генотипа, как например группы крови, цвет глаз и волос, диактилоскопические узоры. Для множества других вопрос может быть решен только в связи с вполне конкретными условиями среды, возрастом и т. д. В связи со сказанным нам надо теперь обратиться к рассмотрению понятия среды и дать критику постулата об «одинаковой» среде. Понятие среды разными исследователями толкуется не всегда вполне одинаково. Логически правильно будет под термином «среда» понимать всю совокупность факторов ненаследственных, т. о. не входящих в понятие генотипа. В таком случае в понятие среды войдут все факторы, действующие вне и внутри организмов матери и отца будущего ребенка и влияющие на гаметы до оплодотворения (питание, болезни, возраст и т. д.); далее, все факторы, влияющие внутри организма матери на эмбриона до рождения, т. о. вся система факторов среды пренатального развития, о которых кратко речь была выше и, наконец, вся система факторов среды, действующих на организм после рождения. Сюда относится бесчисленное количество факторов, весь космос, поскольку он действует на организм, от простейших воздействий неорганических факторов, как свет или температура, до сложнейших факторов социальной среды и т. д. Наконец, надо указать еще на так называемую внутреннюю среду, т. е. на ту сложную и еще очень мало изученную совокупность факторов внутри организма (вероятно, главным образом химических), влияющих на реализацию генотипа (ср. Р. Вагнер и Митчелл, 1958). Общеизвестно, что множество свойств организма животных и человека в той или иной мере образуется до рождения, и потому ряд «прирожденных» признаков, оказывающихся у организма при рождении, нельзя отождествлять с наследственными признаками (ср. Gini, 1940). Нельзя поэтому говорить, например, о «наследственном» сифилисе или туберкулезе на том основании, что эти болезни у некоторых детей оказываются прирожденными и т. д. Методологически очень важным является как можно точнее и глубже выяснить те факторы, которые вызывают возникновение различных прирожденных признаков или являются стимулами для возникновения и развития признаков на разных этапах онтогенеза, в частности в постнатальный период. В этом отношении очень поучителен одни опыт с тритонами (Buchner, 1952). При недостатке кислорода во время развития яиц тритона процесс развития зародышей задерживается, увеличивается жаберный аппарат и возникают различные аномалии мозга, глаз и т. д. как результат задержки развития, т. е. наблюдаются картины, аналогичные описанным на рыбах Стокардом (Stockard, 1921) и другими. Однако если яйца тритона перенести из воды, бедной кислородом, в нормальную воду еще в конце гаструляции, т. е. на столь ранней стадии развития, когда о закладке жаберной системы еще не может быть и речи, то окажется, что последействие кислородного голодания все же скажется в увеличении жаберного аппарата путем образования четвертой добавочной пары жабер. Каким путем осуществляется такое удивительное последействие, пока остается необъясненным. Но самый факт, по-видимому, сомнений не вызывает и показывает, как поздно может проявиться результат воздействия среды в ранний эмбриональный период, как труден анализ причинных связей в подобных случаях и как осторожно надо утверждать такие связи, чтобы не приписать факторам постнатального развития того, что вызвано не ими, а факторами пренатальной среды. Этот момент, по-видимому, не раз игнорировался некоторыми исследователями, что, конечно, не могло не вести к ошибочным выводам. Теперь мы можем перейти к постулату «одинаковой» среды. О таковой обычно говорят в том случае, когда пара близнецов живет в одной семье, одинаково обеспечена всем необходимым, ни один из близнецов не пользуется особыми привилегиями по сравнению с другим, оба примерно одинаково болеют, несут одинаковую учебную нагрузку и т. д. Однако подобная картина среды может слыть за одинаковую только при достаточно большой степени абстракции от ряда деталей ее и влияния их на близнецов. Для известных целей, например для изучения признаков, мало меняющихся под влиянием среды (как рост, цвет глаз),признание такой среды за одинаковую приемлемо. По в других случаях это недопустимо, например, при изучении психики или физиологии высшей нервной деятельности близнецов. В зависимости от состояния здоровья или характера одного из близнецов, например, один из них будет из казалось бы объективно одинаковых условий среды выбирать одни ее свойства, а другой иные. Так, более болезненный партнер будет чаще уединяться, а другой, наоборот, искать общества людей в том же доме или вне его и т. д.; один будет больше сидеть, другой больше двигаться, играть, заниматься спортом и т. д., словом, каждый будет иметь в пределах этой «общей среды» свою индивидуальную «микросреду» (Zazzo, 1955). Это будет отражаться на психическом и физическом развитии того и другого, выражаться в ряде растущих различий между членами такой пары ОБ. Ясно, что каждый из близнецов благодаря выбору нужных ему условий среды живет в несколько иной среде, и различия среды могут в таких случаях оказывать очень глубокое и значительное воздействие, следовательно, говорить, что это «одинаковая» среда, будет просто ошибочно. Поэтому та среда, которая для изучения роста, развития зубной системы или волосяного покрова может сойти за одинаковую, для изучения развития психики, условных рефлексов, даже мышечной системы, сердца или зрения не может в ряде случаев считаться одинаковой. Изучение взаимоотношений субъекта с окружающей средой в таких случаях должно быть достаточно тонким и тщательным, иначе легко прийти к ошибочным выводам. Выбор среды в соответствии со склонностями может очень резко выступить у пары РБ, различие склонностей у членов которой может оказаться в зависимости от неодинаковой наследственности. В таких случаях надо допускать известную обусловленность выбора среды наследственными тенденциями организма (Zazzo, 1955). Иначе говоря, намечается известная взаимозависимость между факторами наследственности и факторами среды. Взаимоотношение наследственности и среды в таком аспекте еще мало изучено. Постулат «одинаковой» среды, конечно, совершенно не выдерживает критики в тех случаях, где особенно заметно выступают внутрипарные различия близнецов, особенно ОБ, вызванные действием факторов пренатальной среды (см. главу пятую). В некоторых работах прежних лет это обстоятельство недостаточно учитывалось. Разумеется, правильная критика постулата «одинаковой» среды ограничивает применение близнецового метода и не позволяет делать легкие выводы и обобщения относительно «доли» наследственности и среды, как это еще недавно наблюдалось в литературе. Различия среды в свою очередь используются близнецовым методом при изучении пары ОБ путем их «взаимоконтроля» (см. главу одиннадцатую), при изучении близнецов, воспитанных врозь (см. главу тринадцатую «ОБ воспитанные врозь») и т. д. Сходство среды, как говорилось, способствует выработке ряда похожих признаков у близнецов даже в том случае, если их генотип не вполне схож в отношении этих признаков. Таким образом, внутрипарное сходство близнецов не может быть всегда сведено только к наследственному сходству по соответствующему признаку. С другой стороны, внутрипарное фенотипическое различие близнецов при наличии генотипического сходства и даже тождества может зависеть от разных факторов среды, и постнатальных и пренатальных, выяснить которые в ряде случаев нелегко, в связи с чем в некоторых работах пренатальные факторы легко упускались из виду, что приводило к ошибочным выводам. Переходим к третьему постулату близнецового метода, согласно которому развитие признаков организма зависит исключительно от двух систем факторов: наследственности и среды. Близнецовый метод, собственно, заимствовал этот постулат от генетики, он лежит в основе этой науки. Из того определения среды, которое мы делали выше, логически невозможно допустить еще иные факторы иных категорий. Поэтому здесь нет надобности критически рассматривать по существу попытки, ведущие за пределы понятий генетики (ср. Verschuer, 1954). Взаимодействие факторов наследственности и среды протекает во времени, — времени онтогенеза. Последовательность и взаимосвязь его фаз имеет чрезвычайно важное значение для понимания взаимодействия факторов наследственности и среды. Генотип, как известно, реализуется в зависимости от возраста, факторы среды по-разному влияют на организм, также в зависимости от возраста. Один год в возрасте 2—3 лет имеет для организма в этом смысле совсем иное значение, чем в возрасте 40—41 года и т. д. Даже такие, казалось бы, стабильные признаки, как, например, форма черепа, в первые годы жизни гораздо изменчивее, чем во взрослом возрасте, и различные факторы среды могут деформировать череп в младенческом возрасте очень заметным образом, тогда как во взрослом возрасте это уже невозможно. Взаимодействие факторов наследственности и среды может быть правильно изучено только с учетом фаз онтогенеза, т. е. возраста. Без такого учета близнецовое исследование может легко привести к ошибочным выводам. Эта сторона дела нередко недооценивалась в ранних работах о близнецах, и пары разного возраста рассматривались как однородный материал, что для ряда признаков, в которых возраст играет существенную роль, недопустимо. Вообще взаимодействие факторов наследственности и среды в связи с процессом онтогенеза еще очень мало исследовано, но представляет огромный интерес и делает близнецов исключительно ценным материалом для его разработки. Разумеется, близнецовый метод не является единственным способом изучения этого и ряда других вопросов, важных для генетики и биологии вообще. Наряду с ним, с привлечением того же материала, но рассмотренного с методически другой точки зрения, может быть использован родословный метод и другие методы генетики, дополняющие друг друга. Из всего вышесказанного видно, что близнецовый метод, критически использованный, есть несомненно ценный метод генетики и в будущем может сыграть немаловажную роль. Им, конечно, не решается в целом проблема соотношения наследственности—среды, как ожидал Гальтон, так как, очевидно, нет единого универсального решения этого вопроса (ср. Newman et al., 1937). Мы видели также, что изучение близнецов не показало, что наследственность всегда играет ведущую роль, являясь своего рода фатумом. Этот устаревший предрассудок, впрочем, давно уже опровергнут и другими генетическими методами.
ПРИЛОЖЕНИЕ. БЛИЗНЕЦЫ У РАСТЕНИИ
Понятия двух основных типов близнецов — ОБ и РБ — возникли и сложились в связи с изучением многоплодия у животных и человека. Однако, как мы увидим далее, эти понятия имеют общебиологическое значение и применимы ко всем организмам, в том числе и к растениям, у которых можно установить половое размножение, с образованием яйцеклетки и зиготы. У растений, в связи с большим многообразием бесполого размножения у них, имеется ряд своеобразных способов возникновения зародышей, которых принимают за близнецов. В этих случаях зародыши возникают не из яйцеклетки, а из других клеток, в той или иной мере по своей природе родственных яйцеклеткам (синергиды, антиподы и др.). Этим растения отличаются от основной массы животных, особенно высших, где такие явления не встречаются. Возникновение двух или больше зародышей в одном семени, обычно называемое в ботанике полиэмбрионией, известно уже давно. Еще Левенгук в 1719 г. описал двух зародышей в семени апельсина. Первая сводка по вопросу о полиэмбрионии была составлена А. Брауном в 1859 г. Страсбургер (Strasburger, 1878) изучал происхождение близнецов у растений вне зародышевого мешка. В дальнейшем полиэмбрионии было посвящено множество работ, обзор которых можно найти в сводках последних лет (Johansen, 1951; Lebegue, 1952; Маheshwari, 1952; Модилевский, 1953; Wardlaw, 1955; Баранов, 1955; Яковлев, 1957, и др.). Мы, конечно, не можем входить здесь в подробности вопроса полиэмбрионии у растений и ограничимся рассмотрением его главным образом с точки зрения общебиологической. Прежде всего нам надо выяснить, в связи с общебиологическими понятиями ОБ и РБ, что же в ботанике называется полиэмбрионией. В мире растений мы не можем весь обширны и круг явлений полиэмбрионии исчерпать понятием ОБ и РБ. Термин «полиэмбриония» употребляется в ботанике в более широком смысле и несколько ином, чем в зоологии. В последней этим словом обозначают явление возникновения двух и более зародышей из одного яйца, т. е. термин «полиэмбриония» по смыслу совпадает с понятием ОБ, и потому РБ уже не подходят под понятие полиэмбрионии в зоологии. В ботанике же этим термином обозначается все огромное многообразие явлений двух и более зародышей в одном семени, независимо от способов их происхождения. Применяя понятие ОБ в ботанике, мы, поэтому, не можем, во-первых, отождествить его с понятием полиэмбрио-нии, так как последнее в ботанике гораздо шире, а во-вторых, не можем понятие ОБ противопоставить понятию РБ и тем исчерпать весь круг явлений близнечества. В ботанике понятию ОБ надо противопоставить понятие не-ОБ, которое должно охватывать, во-первых, РБ, встречающихся у растении, и, во-вторых, все случаи полиэмбрионии не-РБ, куда относятся зародыши, возникшие не из яйцеклетки, а из других клеток: синергид, антипод, нуцеллуса и т. д. Собственно, все эти случаи полиэмбрионии не из яйца не следует считать явлениями близнечества в точном смысле слова. В новейшей ботанической литературе такая точка зрения, отчасти противоречащая общепринятой, уже высказывалась. Так, например, Вардло (Wardlow, 1955) в своей сводке по эмбриологии растений различает «простую полиэмбрионию», которой он обозначает явление образования нескольких зародышей из нескольких оплодотворенных яиц, иначе говоря — РБ, и «полиэмбрионию расщепления» («cleavage polyembryony») — продольное расщепление и разделение на несколько эмбрионов первоначально одного эмбриона (ргоembrio), — очевидно, здесь имеются в виду ОБ. При такой классификации полиэмбрионии вне этого понятия остаются все случаи, обычно относимые сюда в ботанике, т. е. случаи возникновения зародыша не из яйцеклетки, а из иной клетки. Классификация этих явлений у Вардло мало разработана. Иогансен (Johansen, 1951) предлагает словом «полиэмбриония» обозначать только ОБ, т. е. употреблять этот термин в том же смысле, как в зоологии. Однако эти попытки ограничить значение термина «полиэмбриония» еще не являются общепризнанными в ботанике. Так, недавно Лебэг (Lebegue, 1952), придерживаясь немного измененной классификации полиэмбрионии Эрнста (Ernst, 1901), называет «настоящей полиэмбрионией» не только ОБ и РБ, но также все случаи образования зародышей из клеток зародышевого мешка (синергиды, антиподы и т. д.) и даже клеток нуцеллуса — центральной части семяпочки и ее внутренней оболочки, которые он считает приближающимися к почкованию. Лебэг «истинной полиэмбрионии» противопоставляет «ложную полиэмбрионию», когда несколько зародышей возникает из нескольких зародышевых мешков, срастаются два нуцеллуса и т. д. — явления уже далеко стоящие от подлинного близнечества. Понимание термина «полиэмбриония» последнее время не раз обсуждалось в литературе, причем высказывались противоречивые точки зрения (ср. Баранов, 1955, и др.). Предоставляя ботаникам договариваться об объеме понятия полиэмбрионии, мы можем здесь лишь указать, что с точки зрения общей биологии попытки Иогансена и Вардло ограничить понятие полиэмбрионии и приблизить его содержание к принятому в зоологии можно, несомненно, приветствовать. Наконец, нельзя не отметить, что есть ботаники, которые вообще предпочитают термин «близнецы» термину «полиэмбриония» (Fischer, 1956). Переходим теперь к вопросу о способах возникновения ОБ у высших растений, у которых этот вопрос подробнее всего изучен. По-видимому, самым распространенным способом возникновения ОБ является следующий: зигота, дробясь, превращается в многоклеточную массу, которая образует выросты, становящиеся отдельными зародышами. Таким образом получаются ОБ, например, у орхидеи эвлофеи (Eulophea epidendraea, — рис. 195), у некоторых лилейных и других растений. Этот способ напоминает образование ОБ у южных армадилов (см. главу восьмую «Млекопитающие»). В редких случаях наблюдалось раздвоение зиготы на самой ранней стадии дробления: первые две клетки, на которые делится зигота, начиная развитие, могут разойтись и далее развиваться независимо друг от друга, образуя два зародыша или раздвоение верхушечной клетки; например, это наблюдается у одной орхидеи (Cymbidium bicolor). Образование ОБ из первых двух бластомеров известно у морских ежей и других животных (см. главу восьмую «Беспозвоночные»).
Раннее раздвоение зиготы описано, например, у льна (Карpert, 1933); это ясно видно на препаратах. В некоторых случаях у льна обнаружены соединенные близнецы (рис. 196), которые толкуются, как случаи неполного раздвоения первоначально единого зародыша, т. е. так же, как объясняют некоторых соединенных близнецов у животных и человека. Соединенные близнецы описаны также у других растений, например у одного вида перца (Capsicum brutescens), лилии и маиса (Morgan a. Rappleye, 1950, 1951).
В редких случаях наблюдалось раздвоение зиготы на самой ранней стадии дробления: первые две клетки, на которые делится зигота, начиная развитие, могут разойтись и далее развиваться независимо друг от друга, образуя два зародыша или раздвоение верхушечной клетки; например, это наблюдается у одной орхидеи (Cymbidium bicolor). Образование ОБ из первых двух бластомеров известно у морских ежей и других животных (см. главу восьмую «Беспозвоночные»).
Раннее раздвоение зиготы описано, например, у льна (Карpert, 1933); это ясно видно на препаратах. В некоторых случаях у льна обнаружены соединенные близнецы (рис. 196), которые толкуются, как случаи неполного раздвоения первоначально единого зародыша, т. е. так же, как объясняют некоторых соединенных близнецов у животных и человека. Соединенные близнецы описаны также у других растений, например у одного вида перца (Capsicum brutescens), лилии и маиса (Morgan a. Rappleye, 1950, 1951).
 Иногда второй дополнительный зародыш может возникать наподобие почки на первом зародыше; так происходит, например, у махорки (Nicotiana rustica, — Cooper, 1943).
Не останавливаясь на РБ у растений, которые сравнительно еще мало изучены, мы рассмотрим явление полиэмбрионии типа не-РБ, характерное для растений. Чтобы разобраться в этом явлении, надо вспомнить строение зародышевого мешка. Как известно, семя развивается из семяпочки, являющейся макроспорангием; семяпочка состоит из центральной многоклеточной части — нуцеллуса и его покровов — интегумента. В нуцеллусе одна из клеток делится дважды с редукцией числа хромосом. Из получившихся четырех клеток одна сильно разрастается, вытесняя остальные три, — это макроспора или зародышевый мешок. Последний, прорастая, т. е. развиваясь в многоклеточное образование (половое поколение цветковых растений, соответствующее заростку папоротника), вырабатывает яйцеклетку. Рядом с ней располагаются две вспомогательные клетки — синергиды, а на противоположном конце зародышевого мешка — три клетки, называемые антиподами. Вот эти-то клетки, синергиды и антиподы, как уже упоминалось, могут быть оплодотворены и превратиться также в зародышей. Это явление подробно изучено у некоторых растений, например, у лука (Третьяков, 1895; Haberland, 1923, 1925; Модилевский, 1953). Если второй зародыш разовьется из одной из антипод, то он окажется расположенным на противоположном конце зародышевого мешка (рис. 197).
Иногда второй дополнительный зародыш может возникать наподобие почки на первом зародыше; так происходит, например, у махорки (Nicotiana rustica, — Cooper, 1943).
Не останавливаясь на РБ у растений, которые сравнительно еще мало изучены, мы рассмотрим явление полиэмбрионии типа не-РБ, характерное для растений. Чтобы разобраться в этом явлении, надо вспомнить строение зародышевого мешка. Как известно, семя развивается из семяпочки, являющейся макроспорангием; семяпочка состоит из центральной многоклеточной части — нуцеллуса и его покровов — интегумента. В нуцеллусе одна из клеток делится дважды с редукцией числа хромосом. Из получившихся четырех клеток одна сильно разрастается, вытесняя остальные три, — это макроспора или зародышевый мешок. Последний, прорастая, т. е. развиваясь в многоклеточное образование (половое поколение цветковых растений, соответствующее заростку папоротника), вырабатывает яйцеклетку. Рядом с ней располагаются две вспомогательные клетки — синергиды, а на противоположном конце зародышевого мешка — три клетки, называемые антиподами. Вот эти-то клетки, синергиды и антиподы, как уже упоминалось, могут быть оплодотворены и превратиться также в зародышей. Это явление подробно изучено у некоторых растений, например, у лука (Третьяков, 1895; Haberland, 1923, 1925; Модилевский, 1953). Если второй зародыш разовьется из одной из антипод, то он окажется расположенным на противоположном конце зародышевого мешка (рис. 197).
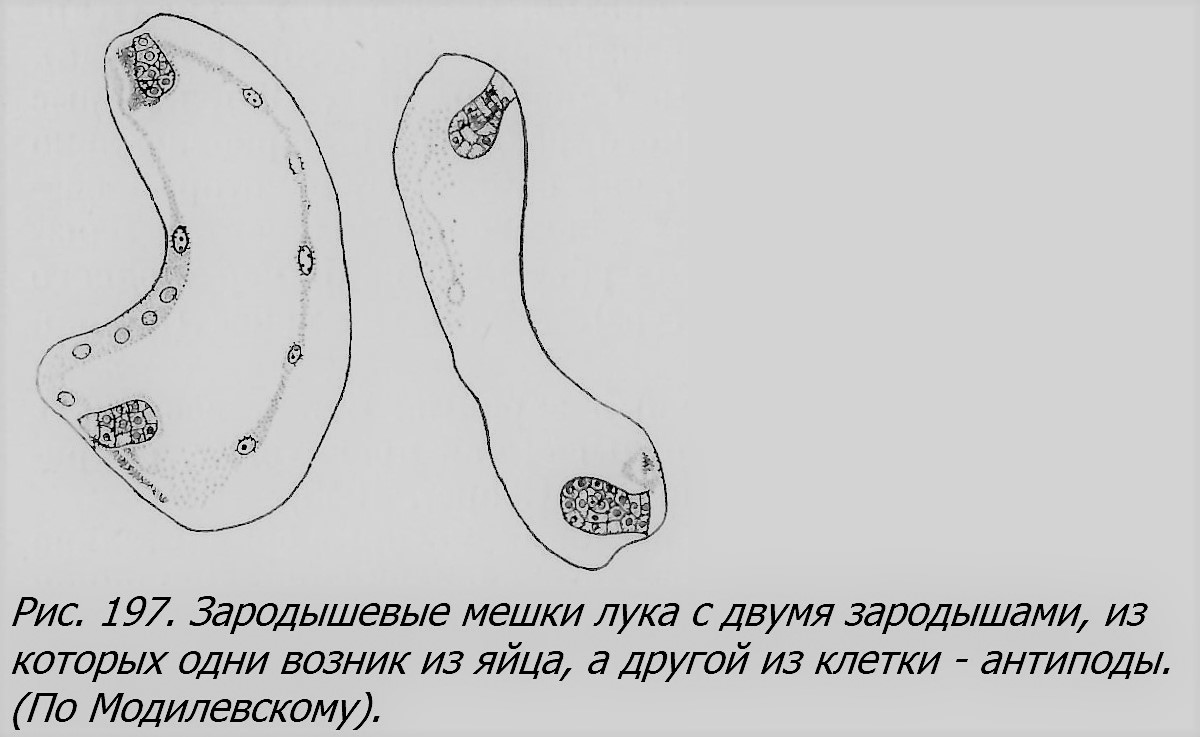 В некоторых случаях встречаются зародыши с гаплоидным (половинным) набором хромосом. Это объясняется тем, что они возникли без оплодотворения, «по индукции», в связи с развитием зиготного зародыша. Обычно в семени выживает только этот зародыш, возникший из оплодотворенного яйца, а прочие зародыши гибнут от истощения (у них нет эндосперма). Мы не будем останавливаться на рассмотрении других случаев полиэмбрионии у растений, в той или иной степени далеких от подлинных явлений близнечества. Они могут дать интересный материал для эволюционного понимания происхождения разных форм полиэмбрионии, вопроса пока еще мало разработанного. Изучение полиэмбрионии у хвойных, у которых она широко распространена, привело, например, Бухгольца (Buchholz, 1933) к предположению, что раньше возникла простейшая форма расщепления зиготы, т. е. образование ОБ, а другие формы полиэмбрионии — позже.
У растений могут возникать не только двойни, но также тройни и четверни. Подробное изучение этого вопроса у спаржи показало, что из 405 случаев полиэмбрионии 97% составляли двойни, 11% — тройни, четверней же был только один случай. Среди близнецов большинство было диплоидных, но были и гаплоидные, триплоидные и другие формы, отличающиеся по набору хромосом (Randall a. Rick, 1945).
Значительный интерес представляет вопрос, чем вызывается образование близнецов. Изучение разных линий у спаржи показало, что самая бедная близнецами линия имеет их 0.13%, а самая богатая ими — 3.5% (Randall a. Rick, 1945). У разных сортов хлопка найдена аналогичная картина (Silow u. Stephens, 1944) и т. д. Такие факты предположительно можно объяснять различием наследственности разных линий в смысле тенденции образовывать близнецов. У льна, например, происходит преимущественное возникновение ОБ путем расщепления молодого эмбриона (Kappert, 1933), и такую тенденцию к раздвоению зародыша считают наследственной. Физиологический механизм этого явления еще мало понятен. Несомненно, что существенное влияние оказывают на это явление факторы среды, например количество воды, содержание калия, время посева и т. д. (Wricke, 1954). В связи с этим можно путем искусственного воздействия вызывать образование ОБ у растений. Так, например, у твердой пшеницы мелянопус 069 путем нанесения на колос распыленного ростового стимулятора — парахлорфеноксиуксусной кислоты — можно вызвать появление на теле зародыша зачатков новых добавочных зародышей (рис. 198). Как показало микроскопическое исследование, мы имеем здесь дело с своеобразной формой возникновения ОБ. В зависимости от направления роста этих зародышей из семени могут выйти или два независимых друг от друга ростка, если они растут в разные стороны, или же, если они растут в одном направлении, они могут в той или иной мере срастись, образуя побег с двумя колосьями и т. п. (Яковлев и Снегирев, 1954).
В некоторых случаях встречаются зародыши с гаплоидным (половинным) набором хромосом. Это объясняется тем, что они возникли без оплодотворения, «по индукции», в связи с развитием зиготного зародыша. Обычно в семени выживает только этот зародыш, возникший из оплодотворенного яйца, а прочие зародыши гибнут от истощения (у них нет эндосперма). Мы не будем останавливаться на рассмотрении других случаев полиэмбрионии у растений, в той или иной степени далеких от подлинных явлений близнечества. Они могут дать интересный материал для эволюционного понимания происхождения разных форм полиэмбрионии, вопроса пока еще мало разработанного. Изучение полиэмбрионии у хвойных, у которых она широко распространена, привело, например, Бухгольца (Buchholz, 1933) к предположению, что раньше возникла простейшая форма расщепления зиготы, т. е. образование ОБ, а другие формы полиэмбрионии — позже.
У растений могут возникать не только двойни, но также тройни и четверни. Подробное изучение этого вопроса у спаржи показало, что из 405 случаев полиэмбрионии 97% составляли двойни, 11% — тройни, четверней же был только один случай. Среди близнецов большинство было диплоидных, но были и гаплоидные, триплоидные и другие формы, отличающиеся по набору хромосом (Randall a. Rick, 1945).
Значительный интерес представляет вопрос, чем вызывается образование близнецов. Изучение разных линий у спаржи показало, что самая бедная близнецами линия имеет их 0.13%, а самая богатая ими — 3.5% (Randall a. Rick, 1945). У разных сортов хлопка найдена аналогичная картина (Silow u. Stephens, 1944) и т. д. Такие факты предположительно можно объяснять различием наследственности разных линий в смысле тенденции образовывать близнецов. У льна, например, происходит преимущественное возникновение ОБ путем расщепления молодого эмбриона (Kappert, 1933), и такую тенденцию к раздвоению зародыша считают наследственной. Физиологический механизм этого явления еще мало понятен. Несомненно, что существенное влияние оказывают на это явление факторы среды, например количество воды, содержание калия, время посева и т. д. (Wricke, 1954). В связи с этим можно путем искусственного воздействия вызывать образование ОБ у растений. Так, например, у твердой пшеницы мелянопус 069 путем нанесения на колос распыленного ростового стимулятора — парахлорфеноксиуксусной кислоты — можно вызвать появление на теле зародыша зачатков новых добавочных зародышей (рис. 198). Как показало микроскопическое исследование, мы имеем здесь дело с своеобразной формой возникновения ОБ. В зависимости от направления роста этих зародышей из семени могут выйти или два независимых друг от друга ростка, если они растут в разные стороны, или же, если они растут в одном направлении, они могут в той или иной мере срастись, образуя побег с двумя колосьями и т. п. (Яковлев и Снегирев, 1954).
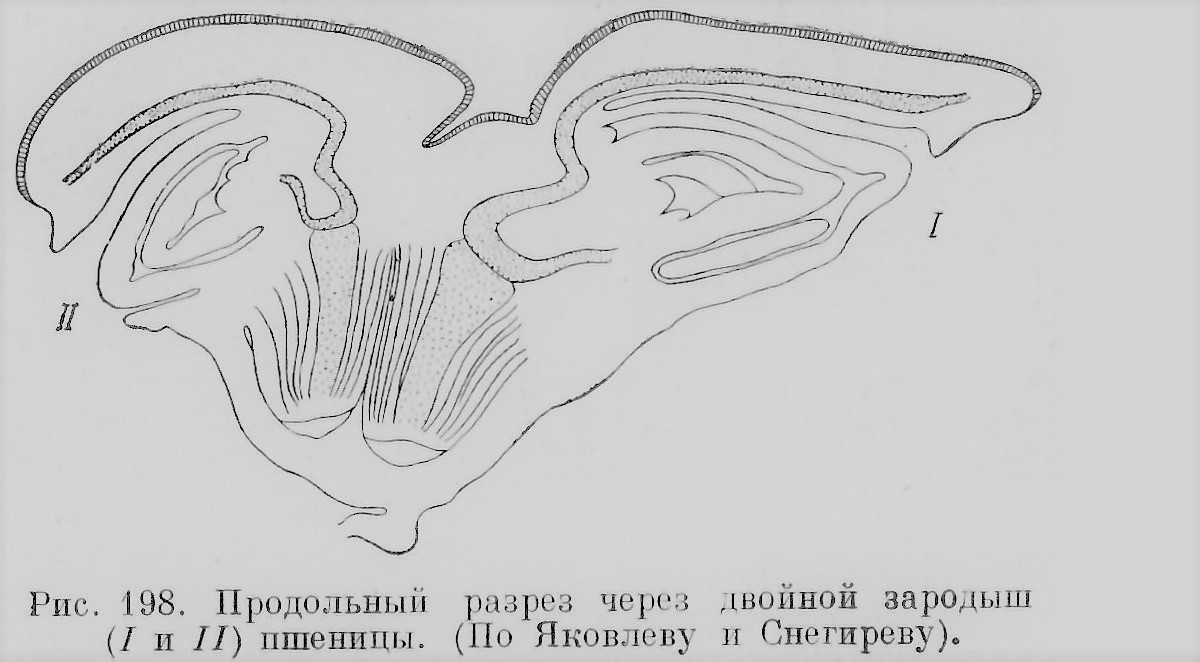 Другие примеры экспериментального получения ОБ могут дать недавно опубликованные опыты Хаккиус (Haccius, 1955), сделанные на весеннике из семейства лютиковых. У этого растения близнецы редки — около 0.03%. При действии на его семена слабым раствором дихлорфеноксиуксусной кислоты, а также других производных уксусной кислоты, удалось вызвать значительное увеличение числа близнецов: 3—8%. Близнецы были однояйцевые, по-видимому, возникшие путем расщепления первоначально одного зародыша. Его нераздвоенный нижний остаток виден на рис, 199 Б, В, обозначенный буквой б. Близнецы возникают вместо семядолей нормального растения. Развитие близнеца на месте одной из семядолей тормозит развитие другой. Основываясь на теории физиологического градиента Чайлда, автор объясняет возникновение ОБ в данных опытах следующим образом: производные уксусной кислоты поражают вершинные клетки роста, как самые активные, по Чайлду, и потому самые чувствительные к вредоносным факторам. Поэтому из клеток, которые нормально образуют семядоли, возникают новые вершинные клетки, образующие близнецовые побеги. Это гипотетическое объяснение образования ОБ у растений вполне аналогично объяснению возникновения ОБ у животных, когда за основу берется теория Чайлда (см. главу четвертую). Насколько это верно и применимо к образованию близнецов в естественных условиях, покажут будущие исследования.
Другие примеры экспериментального получения ОБ могут дать недавно опубликованные опыты Хаккиус (Haccius, 1955), сделанные на весеннике из семейства лютиковых. У этого растения близнецы редки — около 0.03%. При действии на его семена слабым раствором дихлорфеноксиуксусной кислоты, а также других производных уксусной кислоты, удалось вызвать значительное увеличение числа близнецов: 3—8%. Близнецы были однояйцевые, по-видимому, возникшие путем расщепления первоначально одного зародыша. Его нераздвоенный нижний остаток виден на рис, 199 Б, В, обозначенный буквой б. Близнецы возникают вместо семядолей нормального растения. Развитие близнеца на месте одной из семядолей тормозит развитие другой. Основываясь на теории физиологического градиента Чайлда, автор объясняет возникновение ОБ в данных опытах следующим образом: производные уксусной кислоты поражают вершинные клетки роста, как самые активные, по Чайлду, и потому самые чувствительные к вредоносным факторам. Поэтому из клеток, которые нормально образуют семядоли, возникают новые вершинные клетки, образующие близнецовые побеги. Это гипотетическое объяснение образования ОБ у растений вполне аналогично объяснению возникновения ОБ у животных, когда за основу берется теория Чайлда (см. главу четвертую). Насколько это верно и применимо к образованию близнецов в естественных условиях, покажут будущие исследования.
 Получение у растений ОБ — организмов в основном с одинаковым генотипом — представляет большой интерес для опытов с их изменчивостью под влиянием факторов среды. Одни из близнецов может употребляться для опыта, другой — служить контролем. Работы в этом направлении еще не развернуты должным образом. Такие опыты на животных и людях, как известно, уже удалось ставить с успехом.
Мы не будем здесь останавливаться на исследованиях удвоения частей растения, например цветка у настурции (Eyster a. Burpee, 1936) или плода у персика (Евреинов, 1933), у грецкого ореха (Щепотьев, 1950) и т. д. Эти явления, несомненно родственные по природе близнечеству, и до некоторой степени аналогичные явлениям раздвоения конечностей у амфибий и подобным явлениям у животных, касаются не всего организма, лишь части его, и потому лежат уже вне нашей основной задачи.
Получение у растений ОБ — организмов в основном с одинаковым генотипом — представляет большой интерес для опытов с их изменчивостью под влиянием факторов среды. Одни из близнецов может употребляться для опыта, другой — служить контролем. Работы в этом направлении еще не развернуты должным образом. Такие опыты на животных и людях, как известно, уже удалось ставить с успехом.
Мы не будем здесь останавливаться на исследованиях удвоения частей растения, например цветка у настурции (Eyster a. Burpee, 1936) или плода у персика (Евреинов, 1933), у грецкого ореха (Щепотьев, 1950) и т. д. Эти явления, несомненно родственные по природе близнечеству, и до некоторой степени аналогичные явлениям раздвоения конечностей у амфибий и подобным явлениям у животных, касаются не всего организма, лишь части его, и потому лежат уже вне нашей основной задачи.
ЛИТЕРАТУРА[17]
Алекперов А. М. 1954. О нахождении двухголовой змеи. Зоолог, журн., 33 : 716—717. Алексеева Т. Т. 1941. Материалы к проблеме сна (исследования на торакопагах). 9-е совещ, но физиолог. проблемам, Экспер. часть, Тезисы докладов, М.—Л. : 7—8. Алексеева Т. Т. и Н. И. Островская. 1953. Регуляция некоторых физиологических функции на примере сросшихся близнецов. 16-е совещ. по проблемам высш. нервн. деят., М.—Л. : 13—14. Алпатов В. В. 1957. Левизна-правизна в строении растительных и животных организмов. Бюлл. Моск. общ. испыт. прир.,62, 5 : 19—27. Андерсен-Нексе М. 1933. Последыши. (1902). Избранные новеллы. М. : 15—33. (Анохин П. К.) Anokhine Р. 1939. Les jumneaux coalescents. Presse Med., 20 : 379—380. (Ардашников С. H., Е. А. Лихтенштейн, Р. П. Мартынова, Г. В. Соболева и Е. Н. Постникова.) Агdashnikov S. N., Е. A. Lichtenstein, R. Р. Магtуnоva, G. V. Soboleva а. Е. N. Postnikova. 1936 а. The diagnosis of zygosity in twins. J. Hered., 27 : 465—468. Te же. 1936 б. К вопросу о диагностике яйцевости близнецов. Тр. Мед.-генет. инст., 4 : 254—273. Аристотель. О возникновении животных. Кн. 4. Перев. В. Карпова. Изд. АН СССР, 1940 : 173 и сл. Афанасьев А. Н. 1957. Два Ивана солдатских сына. Народные русские сказки. Т. 1. М. : 349—357. Баранов П. А. 1955. История эмбриологии растений. Изд. АН СССР. М.—Л. : 1—439. Барт Л. 1951. Эмбриология. Изд. иностр. лит. : 1—233. Батуев Н. А. 1906. Восемь случаев двойного уродства у человека. Изд. Акад. наук, XXII : 3. Беркович И. М. 1930. Исследование физиологии и патологии близнецов. Сообщ. I. Мед.-биолог. журн., 4—5:288—306. Бляхер Л. Я. 1955. История эмбриологии в России. М. : I—380. Боричевский И. 1840. Повести и предания народов славянского племени. СПб. : 112-116. Босик Л. Я. 1934. К вопросу о роли наследственности и среды в физиологии и патологии детского возраста. Тр. Мед.-биолог, инст., 3 : 33—56. Босик Л. Я., Е. И. Пасынков и И. Б. Гуревич. 1934. Терапевтические исследования на однояйцевых близнецах. Тр. Мед.-биолог. инст., 3 : 119—132. (Босик Л. Я. и Е. И. Пасынков) Воssik L. u. E. Passynkоff. 1936. Therapeutische Forschungen an eineiigen Zwillingen. Behandlung der Rachitis und der Tuberkulosen Bronchadenitis mit Quarzlampenlicht. Monatschr. f. Kinderheilk., 66 : 352—363. Брайант A. T. 1953. Зулусский народ до прихода европейцев. Изд. иностр. лит. (О близнецах : 373—375). Бунак В. В. 1926. О морфологических особенностях одно- и двуяйцевых близнецов. Русск. евген. журн., IV : 23—51. (Бунак В. В.) Bunak V. V. 1927. Particularites morphologiques des jumeaux univilellins et bivitellins. Inst. Int. d’Anthrop., III Sess., Amsterdam : 157. Бунак В. В. 1936. Роль наследственности и среды в изменчивости структуры кожных капилляров (исследование 91 пары близнецов). Тр. Мед.-генет. инст., 4 : 383—400. Бунак В. В. 1941. Антропометрия. Учпедгиз, М. : 1—368. Буханов Я. Г. 1932. К вопросу о групповой характеристике крови двоен. Гинек. и акуш., Год 11-й, 4 : 61—64. Быстрицкий И. А. 1941. Группы крови у близнецов. Тр. Куйбыш. военно-мед. акад., 5 : 285—289. Быховский А. М. 1927. Уроды и уродства. Гл. V. Двойные и множественные уродства. Изд. «Моск. рабочий» : 1—115. (Бэр К. М.) Baer К. 1827. Ueber einen Doppel-Embryo vom Huhne aus dem Anfange des dritten Tages der Bebrutung. Arch. f. Anat. u. Physiol. (Meckel’s Arch.),.. 2 : 476. (Бэр К. M.) Baer К. 1845a. Uber doppelleibige Missgeburten oder organische Verdoppelungen. .Mem. de I’Acad. Sci. St.-Petersb., VI ser., Sci. mathem., phys. et nat., VI, Sec. part., Sci. nat., IV : 79—178. (Бэp К. M.) Baer K. 1845б. Neuer Fall von Zwillingen, die an der Stirnen verwachsen sind. Bull. CI. phys.-math. Acad. Sci. St.-Petersb., III, 8:113—128. (Бэр К. M.) Baer К. 1850. Uber den jetzigen Zustand und die Geschichte des anatomischen Cabinets der Akademie der Wissenschaften zu St.-Petersburg. Сборн. Музея по антрополог. и этногр. Акад. наук, 1900 : 111—152. (Бэр К. М.) Baer К. 1856. Notice sur un monstre double vivant, compose de deux enfants feminins. Bull. CI. phys.-math. Acad. Sci. St.-Petersb., XIV, 1—3 : 34—37. Вагнер P., Г. Митчелл. 1958. Генетика и обмен веществ. Изд. иностр. лит., М. : 1—426. Васильева Г. А. 1956. Редкое двойное уродство. Акуш. и гинек., 6 : 79—80. Вернадский В. И. 1940. Проблемы биогеохимии. IV. О правизне и левизне. М.—Л. : 1—15. Волоцкой М. В. 1937. Близнецовый метод и проблема изменчивости генов. Антропол. журн., 2 : 3—26. Волоцкой М. В. 1936. К вопросу о генетике папиллярных узоров пальцев. Тр. мед.-биол. инст., 4:404—439. (Вольф К. Ф.) Wolff С. F. 1759. Theoria Generationis. Halae. Русск. перев.: Теория зарождения. Изд. АН СССР, 1950 : 1—630. (Вольф К. Ф.) Wolff С. F. 1770. Ovum simplex gemelliferum. Novi Comment. Acad. Sci. Petropol., 14 : 456—483. (Вольф К. Ф.) Wolff C. F. 1773. Descriptio vituli bicipitis cui acced. commentatio de ortu monstrorum. Novi Comment. Acad. Sci. Petropol., 17 : 540—575. (Вольф К. Ф.) Wolff C. F. 1780. Notice touchant un monstre biforme, dont les deux corps sont reunis par derriere. Acta Acad. Sci. Petropol., I, Histoire : 41—44. (Вольф К. Ф.) Wolff C. F. 1783. De pullo monstroso quartuor pedibus totidemque alis instructo. Acta Acad. Sci. Petropol., I : 203— 207. Вольфкович М. И. 1930. Отоларингологическое исследование близнецов. Мед.-биолог. журн., 4—5 : 315—320. Гаузе Г. Ф. 1940. Асимметрия протоплазмы. Изд. АН СССР : 1—128. Гексли Дж. и Г. де Бер. 1936. Основы экспериментальной эмбриологии. Под ред. и дополн. Д. П. Филатова, М.—Л. : 1—468. Гиппократ. О семени и природе ребенка. Избранные книги. Перев. В. И. Руднева. ГИЗ, 1936 : 255 и сл. Груздев В. С. 1914. Уродства и уроды. Библ. «Знание для всех», изд. Сойкина, СПб. : 1—32. Грузова М. Н. 1956. Новые данные но развитию Hydra vulgaris (Pall.). Докл. АН СССР, 109 : 670—672. Гуревич И. Б. 1936. Роль наследственности и среды в изменении размеров сердца. Бюлл. экспер. биолог, и мед., I : 9. Гутскунст В. 1955. Сто тридцать лет дактилоскопии. Przeglad Antropologiszny, 21 : 376. Диллон Я. Г. и И. Б. Гуревич. 1934. Исследование пневматизации придаточных полостей носа и сосцевидного отростка, формы и размеров турецкого седла у близнецов. Тр. Мед.-биолог. инст., 3 : 68—73. 3авадовский М. М. 1941. Гормональный метод стимуляции многоплодия овец. М. : 1—204. Завадовский М. М. 1946а. Активация половой циклики овец в «мертвый» весенне-летний сезон и организация окотов. Вестн. Моск. унив., 2 : 3—24. Завадовский М. М. 1946б. Естественное и экспериментальное многоплодие коров. Вестн. Моск. унив., 3/4 : 97—100. 3авадовский М. М. 1954. Фронтальный вариант гормонального метода стимуляции многоплодия овец. Докл. ВАСХНИЛ, 3 : 20—24. Загорский П. А. 1812. Обозрение разнообразных человеческих уродов. Умозрительные исследования. СПб., 3 : 265—277. Загорский П. А. 1815. Описание и изображение двойни, сросшейся между собой передней частью тела. Умозрительные исследования. СПб.; 4 : 382—399. 3агорский П. А. 1832. Описание и изображение близнецов, лбами между собой сросшихся. В кн.: М. А. Тикотин. П. Загорский. М., 1950 : 253—256. Закиров И. 3. 1956. Случай двухголового уродства. Сборник научн. трудов Самаркандск. мед. инст., 9:305—309. Занд Жорж. (1848). Маленькая Фадетта. Собр. соч., 3, Л., 1929 : 1 —116. Зелинский Ф. Ф. 1922. Плавт и Шекспир. («Менехмы» и «Комедия ошибок»). В кн. : Возрожденцы. I. СПб. : 81 —107. (Злотников М.) Zlоtniкоff M. 1945. A human mosaic. Bilaterally asymmetrical naevus pigmentosus pilosus et mollusciformis unilateralis. J. Hered., 36 : 163—168. Иванов П. П. 1945. Руководство по общей и сравнительной эмбриологии. Л. : 1—351. Иванова Е. М. 1927. Глаза близнецов. Арх. офтальмолог., 3 : 310—314. Игнатьев М. В. 1936. К вопросу о математической интерпретации близнецовых корреляций. Тр. Мед.-генет. инст., 4 : 284—295. Кабаков И. Б. 1950. Анализ электрокардиограмм у близнецов. Мед.-биолог, журн., 4—5:307—314. Кабаков И. Б. и И. А. Рывкин. 1934. Исследование электрокардиограмм у близнецов. Сообщ. II. Роль наследственности и среды в изменчивости электрокардиограммы. Тр. Мед.-биол. инст. 3 : 73—79. Кабаков И. Б., И. А. Рывкин и И. Б. Гуревич. 1934. Исследование электрокардиограмм у близнецов. Сообщ. III. Об изменчивости зубца Т. Тр. Мед.-биол. инст., 3 : 80—85. Канаев И. И. 1934. Опыт изучения условных рефлексов у однояйцевых близнецов. Арх. биолог. наук, 34 : 569—577. Канаев И. И. 1937. Дальнейшее изучение физиологической деятельности мозга у однояйцевых близнецов. Арх. биолог. наук, 44 : 13—42. (Канаев И. И.) Kanajev J. 1938. Physiology of the brain in twins. Character a. personality, 4, 3 : 177—187. Канаев И. И. 1939a. Спонтанная слюнная секреция у человеческих близнецов. Докл. АН СССР, 25 : 252—254. Канаев И. И. 1939б. Безусловные слюнные рефлексы у человеческих близнецов. Докл. АН СССР, 25 : 255—258. Канаев И. И. 1939в. Условные слюнные рефлексы у человеческих близнецов. Докл. АН СССР, 25 : 259—260. Канаев И. И. 1940а. К вопросу о корреляции количественных показателей дактилоскопических рисунков у однояйцевых близнецов. Докл. АН СССР, 26 : 82—84. Канаев И. И. 1940б. Слюнные рефлексы у близнецов. Природа, 11 : 81-85. Канаев И. И. 1941. К вопросу о подвижности условных рефлексов у близнецов. Докл. АН СССР, 30, 9 : 851—853. Канаев И. И. 1948. Экспериментальная генетика высшей нервной деятельности человека. Усп. совр. биолог., 25 : 149—155. Канаев И. И. 1951. Работы К. М. Бэра о соединенных близнецах и о некоторых других аномалиях развития. Природа, 4 : 75—79. Канаев И. И. 1954. О влиянии среды на развитие высшей нервной деятельности. Природа, 4 : 107—110. Канаев И. И. и Н. В. Боровка. 1939. Опыт изучения корреляции между спонтанной, условной и безусловной слюнной секрецией у одной пары однояйцевых близнецов. Докл. АН СССР, 25 : 261 — 262. Канаев И. И. и А. С. Кульминская. 1959. О корреляции между нервными процессами у близнецов. Журн. высш. нервн. деят., 9 : 212—215. Короткин И. И. и Н. А. Крышова. 1948. Изменение моторной хронаксии во время сна у двух однояйцевых близнецов. Физиолог. журн., 34 : 229—232. Крейцвальд Ф. Г. 1953. Сказка о Быстроногом, Ловкоруком и Зоркоглазом. Старинные эстонские народные сказки. Таллин : 17 и сл. Крюи Поль де. 1937. Стоит ли им жить? М. (О близнецах Дионн : 192— 213). Кузнецов Д. И. 1951. Ненормальное развитие цыпленка. Природа, 8 : 77—78. Кушнер X. и Е. Панфилова. 1955. Редкий случай однояйцевой двойни у кур. Журн. общ. биолог., 16 : 248. (Лебединский Н. Г.) Lebedinsky N. G. 1923. Uber eine Duplicitas anterior von Rana fusca und uber die teratogenetische Terminationsperiode der symmetrischen Doppelbildungen der Placentalier. Anat. Anz., 56 : 257—266. (Левит С. T. и Г. В. Соболева) Levit S. T. a. G. V. Soboleva. 1935. Comparative intrapair correlations of fraternal twins and siblings. J. Genet., 30:389—396. Левит С. T. и Г. В. Соболевa. 1936. Сравнительная внутрипарная корреляция двуяйцевых близнецов и братьев-сестер. Невролог. и генет., 1 : 63—70. Лихтенштейн Е. А. и Н. Л. Басина. 1936а. Исследование полости рта у близнецов. Сообщ. I. Кариес. Тр. Мед.-генет. инст., 4 : 305—327. Лихтенштейн Е. А. и Н. Л. Басина. 1936б. Исследование полости рта у близнецов. Сообщ. II. Прорезывание, смена, морфология и расположение зубов. Тр. Мед.-генет. инст., 4 : 328—352. (Лурия А. Р.) Luгiа А. 1936—1937. The development of mental functions in twins. Character a. personality, 5:35—47. Луpия A. P. и A. H. Mиpeновa. 1936a. Исследование экспериментального развития восприятия методом дифференциального обучения однояйцевых близнецов. Невролог. и генет., 1 : 407—443. Лурия А. Р. и А. Н. Миренова. 1936б. Экспериментальное развитие конструктивной деятельности. Дифференциальное обучение однояйцевых близнецов. Тр. Мед.-генет. инст., 4 : 487—505. Лурия А. Р. и Ф. Я. Юдович. 1956. Речь и развитие психических процессов у ребенка. М. : 1—93. Маршак С. 1954. Про Сережу и Петю. Журн. «Огонек», 11 : 28. Малкова Н. Н. 1934. Роль наследственности и среды в изменчивости высоты кровяного давления и частоты пульса. Тр. Мед.-биолог. инст., 3 : 57—68. Мартынова Р. П. 1945. К вопросу о роли наследственности в этиологии рака. Бюлл. экспер. биолог, и мед., 19, 3 : 12 —15. Мартычьянц А. И. 1932. Урологическое обследование близнецов. Урология, 9 : 35—38. Менделевич Э. К. 1936. Наблюдения над инфекционными заболеваниями у близнецов. Бюлл. экспер. биолог. и мед., 2 : 248—250. Миллер Вс. Ф. 1876. Очерки арийской мифологии в связи с древнейшей культурой. Т. I. Асвины-Диоскуры. М. : 1—355. Миренова А. Н. 1932. Обучение и рост у однояйцевых близнецов. Психология, 4 : 119—123. Миренова А. Н. 1934. Психомоторное обучение дошкольника и общее развитие. Некоторые эксперименты на близнецах. Тр. Мед. биол. инст., 3 : 86—103. Миренова А. Н. и В. Н. Колбановский. 1934. Сравнительная оценка методов развития комбинаторных функций у дошкольника. Эксперименты на однояйцевых близнецах. Тр. Мед.-биол. инст., 3 : 104—118. (Миренова А. Н.) Mirenova А. N. 1935. Psychomotor education and general development of pre-school children. Experiments with twin controls. Genet. Psychol., 46 : 433—454. Митрофанов П. 1892. Исследования над развитием позвоночных животных. Работы зоотом. лабор. Варшавск. унив., 1 : 1—251. (Митрофанов П.) Мitrophanow Р. 1895. Teratologische Studien. Arch. Entw.-Mech., 1. Митрофанов П. И. 1899. Тератологические наблюдения. Новая серия. Работы зоотом. лабор. Варшавск. унив., 22 : 1—64. (Митрофанов П. И.) Mitrophanow Р. I. 1899. Notes embryologiques et teratogeniques. C. R. Assoc. Anat., I sess., Paris : 87—99. Модилевский Я. С. 1953. Эмбриология покрытосемянных растений. Киев : 173—177. Молчанова-Сeничева Е. Н. 1941. Условные рефлексы у однояйцевых близнецов на первых месяцах жизни. (Предв. сообщ.). 9-е совещ. по физиолог. проблемам, Экспер. часть, Тезисы докладов, М.—Л. : 56—57. Наранович П. 1856. Анатомическое описание близнецов, сросшихся головками. СПб. : 1—16. Николаев Л. П. 1940. Дискордантность некоторых соматологических признаков у ОБ дошкольного и школьного возраста. Педiатр., акуш. i гiнек., 1 : 11—17. Ниль Дж. и У. Шэлл. 1958. Наследственность человека. Изд. иностр. лит., М. (Гл. XVI. Близнецы : 295—319). Петской П. Г. 1953. Понятие и типы эмбрионального парабиоза. Докл. АН СССР, 89 : 693—694. Плавт Т. М. Близнецы (Menaechmi). Избранные комедии. М., 1933 : 87—156. Райков Б. Е. 1952. Русские биологи-эволюционисты до Дарвина. I. М.—Л. : 163 и сл. Рывкин Н. А. 1936. Исследование электрокардиограммы у близнецов. Сообщ. IV. Некоторые корреляции физиологических и патологических особенностей сердца. Тр. Мед.-генет. инст., 4 : 440—465. Соболева Г. В. 1926. Результаты обследования 105 пар близнецов г. Москвы. Русск. евген. журн., IV : 3—22. Соболева Г. В. и М. В. Игнатьев. 1936. О гено- и паратипической обусловленности роста и веса. Тр. Мед.-генет. инст., 4 : 370— 377. Соколов Б. М. 1925. Анатомическое исследование плода-урода типа duplicitas anterior (dicephalus et bicollis). Русск. арх. анат., гистолог. и эмбриолог., 4 : 119—140. Соловьев Ф. А. 1911. Об однояйцевых близнецах. Патолого-анатомическое исследование. Харьков : 1—74. Спирова М. С. 1928. Три случая двойного уродства вида Prosopothoracopagus у человека. Сборн. Музея антрополог. и этногр., 7 : 208—220. Спирова М. С. 1929. Четыре случая двойного уродства ileothoracopagus monosymmetros у человека. (По коллекции Антропологич. отд. МАЭ). Сборн. Музея антрополог. и этногр., 8 : 215—224. Станюкович Т. В. 1953. Кунсткамера Петербургской Академии наук. М.—Л. : 1—240. Суханов С. А. 1900. О психозах у близнецов. Клинич. журн., 4 : 341—352. Тарецкий А. 1880. Двойные уроды с разделением нижней половины тела. Воен.-мед. журн., 4, 137 : 219—255. Твэн Марк. Избранные рассказы. М., 1936. (Визит интервьюера: 127, Сиамские близнецы : 159—163). (Тонков В. Н.) Тоnкоff W. 1900. Experimentelle Erzeugung von Doppelbildungen bei Triton. Sitzungsber. Berl Akad. Wissensch. : 794—797. Тонков В. Н. 1904. Развитие двойных уродств из нормального яйца. Тр. СПб. общ. естествоисп., 35, 3 : 82—83. (Третьяков С.) Tretjakow S. 1895. Die Beteilung der Antipoden in Fallen der Polyembryonie bei Allium odorum. Ber. Deutsch. bot. Gesellsch., 13, 13. Tуp И. Я. 1903. К казуистике и теории многозачатковых уродств. Работы зоотом. лаб. Варшавск. унив., 29. (Тур И. Я.) Тur J. 1903. Sur un cas de diplogenese tres jeune dans le blastoderme de Lacerta occellata. Bibliograph. Anat.,12, 3 : 83—88. Tуp И. Я. 1904a. К вопросу об эмбриональных включениях («Foetus in foetu»). Варшавск. унив. изв., II : 1—11. (Тур И. Я.) Тur J. 1904. Contributions a la theorie des polygeneses. C. R. Soc. Biol. Paris, 56 : 108—110. (Tур И. Я.) Тur J. 1913. Sur les diplogeneses embryonnaire a centres rapproches. Arch. Biol., 28 : 325—345. Tуp И. Я. 1914. Исследования над развитием сложных уродств. Варшавск. унив. изв., IV : 1—93. Тургенев И. С. 1878. Близнецы. Стихотворения в прозе. Собр. соч., 8, М., 1956 : 511. Цвейг С. Легенда о сестрах-близнецах. Избр. соч., I, М., 1956 : 445—461. (Чурилов М.) Tchouriloff М. 1878. Sur la statistique des naissances gemellaires et leur rapport avec la taille. Bull. Soc. anthropol. Paris, 12, II ser., annee 1877 : 440—446. Шевченко Тарас. (1855—1856). Близнецы. Собр. соч., 4, М., 1949 : 7—139. Шекспир В. Комедия ошибок. Полн. собр. соч., 2, М., 1958 : 103— 294. СПб. Шекспир В. Двенадцатая ночь или что угодно. Избр. произв., М,—Л., 1950 : 352—383. Шимкевич В. М. (год не указан). Уродства и происхождение видов. Изд. Вольфа, Пгр. : 1—106. (Шмидт Г. A.) Schmidt G. А. 1933. Schnurungs- und durchschneidungsversuche am Anurenkeim. Roux’Arch., 129 : 1—44. Шмидт Г. A. 1937. Исследования над регуляцией осевых органов у бесхвостых амфибий. 1. Регуляция осевых органов в стадии бластулы. Биолог. журн., 6 : 513—564. Штернберг Л. Я. 1916. Античный культ близнецов в свете этнографии. Сборн. Музея антрополог. и этногр. Акад. наук, 3. Также в кн.: Первобытные религии в свете этнографии. Л., 1936 : 72—108. (Нем. перевод в: Ztschr. Ethnol., 61. Jg. 1/3, 1930). Штернберг Л. Я. 1925. Лекции по эволюции религиозных верований. Лекц. XVI. Культ. близнецов. В кн.: Первобытная религия. Л.. 1936 : 360—365. Штернберг Л. Я. 1927. Культ близнецов в Китае и индийские влияния. Сборн. Музея антропол. и этногр., 6. Также в кн. : Первобытная религия в свете этнографии. Л., 1936 : 127—140. Щепотьев Ф. Л. 1950. Плоды-близнецы грецкого ореха. Природа, 11 : 56—59. Эфраимсон В. П. 1957. Основные данные современной генетики и действие ионизирующей радиации на наследственные факторы. Бюлл. Моск. общ. испыт. прир., отд. биолог., 6 : 5 — 18. Юдин Т. И. 1907. Психозы у близнецов. Журн. невропатол. и психиатр. им. Корсакова, VII, 1 : 68—83. Юдин Т. И. 1924. Сходство близнецов и его значение в изучении наследственности. Русск. евген. журн., 11 : 28—49. Яковлев М. С. 1957. Основные типы полиэмбрионии высших растений. Тр. Ботан. инст. им. Комарова АН СССР, Морфолог. и анат. раст., IV : 201—210. Яковлев М. С. и Д. П. Снегирев. 1954. Влияние ростковых веществ на образование многозародышевых зерновок у пшеницы. Ботан, журн., 39 : 187—194. Яцута К. 3. 1912. Систематическое иллюстрированное описание коллекции уродов Музея антропологии и этнографии им. Петра Великого при Академии наук. Вып. I. Симметричные близнецы. Асимметричные близнецы. СПб. : 1—44. Яцута К. 3. 1913. Систематическое иллюстрированное описание коллекции уродов Музея антропологии и этнографии им. Петра Великого. Вып. II. Янусовидные уроды — Cephalothoracopagi : 1—58. Яцута К. 3. 1915. Систематическое иллюстрированное описание коллекции уродов Музея антропологии и этнографии им. Петра Великого. Вып. III. Сросшиеся грудной клеткой — Thoracopagi. Сборн. Музея антропол. и этногр., II, 4, : 101—152.Abel W. 1932. Physiognomische Studien an Zwillinge. Ztschr. Ethnol., 64 : 379—388. Ahlfeld F. 1880—1882. Die Missbildungen des Menschen. Leipzig. Aird J. 1954. The conjoined twins of Kano. Brit. Med. Journ., 4866 : 831—837. Allee W. C. 1951. Cooperation among animals. With human implications. N. Y. : 148—153. Allen G. 1955a. Differences between fraternal twins. J. Hered., 46 : 262. Allen G. 1955b. Comments on the analysis of twin samples. Acta Gen. Med. Gemellolog., IV : 143—159. Allen G. a. F. Кallmann. 1955. Frequency and types of mental retardation in twins. Amer. J. Hum. Gen., 7 : 15—20. Alm I. 1953. Differences in the mortality of single and multiple premature births. Acta Paediatr., 42 : 487. Anderson R. E. et al. 1951. The use of skin grafting to distinguish between monozygotic and dizygotic twins in cattle. Heredity, 5 : 379—397. Apert E. 1923. Les jumeaux, etude biologique, physiologique et medicale. Paris. Areу L. B. 1922a. Chorionic fusion and augmented twinning in the human tube. Anat. Rec., 23 : 253—262. Areу L. В. 1922b. Direct proof of the monozygotic origin of human identical twins. Anat. Rec., 23 : 245—251. Areу L. В. 1923. The cause of tubal pregnancy and tubal twinning. Amer. J. Obst. a. Gynec., 5 : 163. Arieti S. 1944. An interpretation of the divergent outcome of Schizophrenia in identical twins. Psychiatr. Quart., 81 : 587—599. Arnold J. 1912. Adult human ovary with follicles containing several oocytes. Anat. Rec., 6 : 413—422. Asshetоn R. 1898. An account of the blastodermic vesicle of the sheep of the seventh day with twin germinal areas. J. Anat. a. Physiol., N. S., 12, 32:362—370. Asshetоn R. 1913. Fisson of the embryonal area in mammals. IX Congr. Intern. Zool. Monaco : 415—422. Вab H. 1906. Geschlechtsleben, Geburt und Missgeburt in der asiatischen Mythologie. Ztsclir. Ethnol., 28. Ballantyne J. H. 1897. Teratogenesis, an inquiry on the causes of monstruosities. History of the past. Edinburgh. Baillarger. 1855. Recherches statistiques, physiologiques et pathologiques sur les enfants jumeaux. C. R. Arad. Sci. Paris. 41 : 931. Barfurth. 1895. Ein Zeugnis fur eine Geburt von Sieblingen beim Menschen. Anat. Anz., 10 : 330—332. Barrer L. 1947. Unilateral Hexadactyly in man. J. Hered., 38 : 345 : 346. Bass M. H. 1952. Diseases of the pregnant woman affecting the offspring. Adv. Int. Med., 5 : 15—58. Bataillоn E. 1901. La pression osmotique et les grands problemes de la biologie. Arch. Entw.-Mech., 11. Baudоuin M. 1901. La vie des monstres doubles; la croissance de Rosa-Josepha etc. Aesculape, 1 : 60—68. Bauer K. 1927. Homoiotransplantation von Epidermis bei eineiigen Zwillinge. Bruns'Beitr. z. klin. Chir., 141 : 442—447. Вecher H. 1935. Anatomische Untersuchungen an eineiigen Zwillingen. Verh. anat. Ges. Jena, 81 : 255. Bell J. 1933. Plural births with a new pedigree. Biometrika, 25 : 110—120. Berry H., Th. Dobzhansky, Garther, Levene, Osborne. 1955. Chromatographic studies on urinarry excretion patterns in monozygotic and dizygotic twins. I—II. Amer. J. Hum. Gen., 7 : 93—107, 108—121. Bertillon L. A. 1874. Des combinaisons de sexe dans les grossesses gemellaires (doubles on triples), de leur causes et de leurs caracteres ethniques. Bull. Soc. Anthrop. Paris, 2 ser., 9 : 270 290. Billingham R. et al. 1952. Tolerance to homografts, twin diagnosis and the freemartin condition in cattle. Heredity, 6 : 201—212. Birkenfeld W. 1932. Beitrag zur Zwillingspathologie der Mamma. Arch. f. klin. Chir., 168 : 568. Bissonnette T. H. 1933. A two-faced kitten. J. Hered., 24 : 103—104. Black H. 1940. Coalescent twins. J. Hered., 31 : 16—18. Вland-Sutton J. 1922. Rosa-Josepha Blazek. the Bohemian twins. Lancet, 1 : 772. Вlatz W. E. 1938. The five sisters. N. Y. Воdemann E. 1935. A case of uniovular twins in the mouse. Anat. Rec., 62 : 291 294. Воlк L. 1910. Die Furchen an den Grosshirnen eines Thorakopagen. Folia neuro-biolog., IV : 207—217. Воlк L. 1926. Die Doppelbildung eines Affen. Beitr. patholog. Anat. u. allg. Pathol., 76 : 238—253. Boltоn G. 1830. Statement of the principal circumstances respecting the united siamese twins now exhibited in London. Phil Trans.: 177—186. Bonnier G. a. A. Hanssоn. 1946. Studies on monozygous cattle twins. V. Acta Agric. Suecana, 1 : 171—205. Bonnier G. a. A. Hanssоn. 1948. Identical twin genetics in cattle. Heredity, 2 : 1—27. Воnnevie K. 1919. Erblichkeit von Zwillingsgeburten Dtsch. med. Wochenschr., 38 : 1059. Воnnevie K. u. A. Sverdrup. 1926. Hereditary predispositions to dizygotic twin-births in Norwegian peasant families. J. Genet., 16 : 125. Воrchardt L. 1931. Intestinaler Infantilisinus und Basedowsche Krankheit als Ursache wesentlicher Verschiedenheiten bei eineiigen Zwillingen. Ztschr. f. Konstitutionsl., 16: 123—129. Born G. 1897. Uber Verwachsungsversuche mit Amphibienlarven. Arch. Entw.-Mech., 4 : 517—623. Bouterwek H. 1943. Erbe und Personlichkeit. Charakterologische Ergebnisse der Zwillingsforschung. Wien. Воvet D. 1931. L’orientation des visceres chez les truites doubles. Bull. Biol. France et Belg., 65 : 216—233. Bowen R. 1953. Allergy in identical twins. J. Allergy, 24, 3 : 236—244. Braam-Houckgeest A. et J. Sanders. 1940. Jumeaux identiques concordants avec nanisme hypophysaire. Genetica, XXII : 261—263. Bracken H. 1936. Uber die Sonderart der subjektiven Welt von Zwillingen. Arch. Ges. Psychol., 97 : 97—105. Bracken H. 1939a. Das Schreibtempo von Zwillingen. Ztschr. menschl. Vererb.- u. Konstitutionsl., 23 : 278—297. Bracken H. 1939b. Wahrnehmungstauschungen und scheinbare Nachbildgrosse bei Zwillingen. Arch. Ges. Psychol., 103 : 203—230. Вrander T. 1937. Uber die Zwillingsforschungen und ihre Beruhrungspunkte mit der Kinderheilkunde. Acta Paediatr., 21 :5—136. Вrander T. 1938. Uber die Bedeutung des Unternormalen Geburtsgewichtes fur die weitere korperliche und geistige Entwicklung der Zwillinge. Ztschr. menschl. Vererb.- u. Konstitutionsl., 21 : 306. Bratengeier E. 1934. Uber die Variabilitat der Frontzahne bei ein- und zweieiigen.Zwillingen. Diss., Frankfurt. Brauer A. 1917. Uber Doppeldungen des Skorpions (Euscorpius carpathicus L.) Sitzungsb. Preuss. Akad. Wiss., Phys.-math. Kl., XII—XIII : 208—221. Brauer A. 1938. Modifications of development in relation to differential susceptibility of the bruchid (Coleoptera) egg to KCN during different metabolic phases. Physiol. Zool., 11, 3 : 249—266. Brauns L. 1934. Studien an Zwillingen im Sauglings- und Kleinkin desalter. Ztschr. f. Kinderforsch., 43 : 86—129. Breakstone B. 1922. The last illness of the Blazek (Grown-together) twins. Amer. Medic., 28, compl. ser., 17 : 221—226. Breitinger E. 1955. Zur Morphogenese und Typologie der Brauen. Homo, 6 : 5—19. Brodnitz Fr. 1951. Stuttering of different types in identical twins. J. Speech. Hearing Disorders, 16 : 334—336. Brody S. 1952. Variation in size and weight of twins of monochorital pregnancies. Amer. J. Obst. Gynecol., 64 : 340—346. Brown J. B. 1937. Homografting of skin: with report of success in identical twins. Surgery, 1 : 558—563. Вruскer W. H. 1944. Influence of heredity and environment on the caries pictures of monozygotic twins. J. Amer. Dent. A., 31 : 931—940. Bruins J. 1955. European congress of twins. Acta Gen. Med. Gemellolog., IV : 365—368. Buchholz J. T. 1933. Determinate cleavage polyembryony with special reference to Dacrydium. Bot. Gaz., 94 : 579. Buchner Fr. 1952. Zur Biologie und Pathologie der Entwicklung. Med. Klin., 47 : 605—611. Burger P. 1935. Jumeaux monoamniotiques. Bull. Soc. obst. gynecol., 24 : 116—118. Burns W. 1949. Hereditary myopia in identical twins. Brit. J. Ophthalm., 33 : 491—494. Buschke F. 1934. Rontgenologische Skelettstudien an menschlichen Zwillingen und Mehrlingen. Leipzig. Buschke F. 1935. The Radiological examination of the skeletons of triplets. J. Hered., 26 : 391—410. Busse H. 1936. Uber normale Asymmetrien des Gesichts und im Korperbau des Menschen. Ztschr. Morph, u. Anthrop., 35 : 412—415. Вutz H. u. Schmahlstieg. 1953. Neuere Ergebnisse der Zwillingsforschung bei Rind und Pferd. Berl. u. Munch, tierarztl. Wochenschr., 66 : 222—225. Byerly T. a. M. Olsen 1934. Polyembryony in the domestic fowl. Science, 80 : 247—248. Campton E. 1926. Partial twinning in pigs. J Hered., 17 : 411—412. Castle W. E. et al. 1936. Identical twins in a mouse cross. Science, 84 : 581. Caullery M. 1945. Biologie des jumeaux. Paris. 168 p. Chapman A. a. J. Lush. 1932. Twinning, sex ratios, and genetic variability in birth weight in sheep. J. Hered., 23 : 473—478. Charache H. 1953. Tumors in one of homologous twins. Amer. J. Roentgenol., 70 : 810—813. Child С. M. 1941. Patterns and problems of development. Chicago. Christensen H. a. R. Вamfоrd. 1943. Haploids in twin seedlings of Pepper capsicum annuum L. J. Hered., 34 : 99—104. Clarke A. a. D. Revell. 1930. Monozygotic triplets in man. J. Hered., 21 : 147—156. Clarke A. a. D. Revell. 1932. Identical quadruplets. J. Hered., 23 : 256. Сlaussen F. 1940. Zur Phanogenese von Gaumenspalten und angeborenen Herzfehlern, ein Beitrag aus der Zwillingskasuistik. Erbarzt, 8 : 5—12. Cleveland M. 1926. Situs inversus viscerum. Arch. Surgery, 13: 343—368. Cock A. G. 1950. A case of incomplete twinning in the rabbit. J. Genet., 5 : 59—66. Cockayne E. 1940. Transposition of the viscera and other reversals of symmetry in monozygotic twins. Biometrica, 31 : 287—294. Cohrs P. 1934. Eineiige Zwillinge bei Schaf und Schwein und zweieiige, aber monofollikulare Zwillinge beim Schaf. Berl. tierarztl. Wochenschr., 39 : 611—645. Coke Fr. 1937. Identical twins. Lancet, 233 : 655. Collier Th 1948. Pseudohermaphroditism in twins (Report of tenth case). Amer. J. Dis. Child.. 76 : 209—219. Colombo R. 1559. Do re anatomica Libri XV. Venetiis. Conklin E. 1933. The development of isolated and partially separated blastomeres of Amphioxus. J. Exp. Zool., 64 : 303—352. Conrad K. 1935—1936. Erbanlage und Epilepsie. Untersuchungen an einer Serie von 253 Zwillingspaaren. Ztschr. Neurol. Psychiatr. 153 : 271—326, 155 : 254—297, 509—542. Cook R. 1934. «Septuplets» with different birthdays. J. Hered., 25: 193—197. Corner G. W. 1922. The morphological theory of monochorionic twins as illustrated by a series of supposed early twin embryos of the pig. J. Hopkins Hosp. Bull., 33 : 389—392. Corner G. 1955 The observed embryology of human single-ovum twins and other multiple births. Amer. J. Obst. Cynecol., 70 : 933— 951. Crabb E. 1931. The origin of independent and of conjoined twins in fresh-water snails. Arch. Entw.-Mech., 124 : 332—356. Cummins H. 1930. Dermatoglyphics in twins of known chorionic history, with reference to diagnosis of the twin varieties. Anat. Rec., 46 : 179—198. Cummins H. 1936. Finger prints in «siamese» twins. Eug. News, 21 : 89—95. Cummins H. 1940. Finger prints correlated with handedness. Amer. J. Phys. Anthrop., 26 : 151—166. Cummins H. 1942. A fraternal twin pregnancy. Amer. J. Obst. Gynecol., 43 : 1041 — 1044. Cummins H. a. G. Mairs. 1934. Finger prints of conjoined twins. J. Hered., 25 : 237 —243. Cummins H. a. Midlo. 1943. Finger prints, palms and soles. Philadelphia : 1—339. Curtius Fr. 1930. Nachgeburtsbefunde bei Zwillingen und Aehnlichkeitsdiagnose. Arch. Gynakol., 104 : 361—366. Curtius Fr. u. G. Коrкhaus. 1930. Klinische Zwillingsstudien. Ztschr. f. Konstitutionsl., 15 : 229—267. Curtius Fr. u. O. Verschuer. 1932. Die Anlage zur Entstehung von Zwillingen und ihre Vererbung. Arch. Rass. u. Gesellsch. Biol., 26 : 361 — 387. Dahlberg G. 1926. Twin births and twins from a hereditary. Point of view. Stockholm : 1—269. Dahlberg G. 1929. Genotypische Asymmetrien. Ztschr. ind. Abstamm.- u. Vererbungsl., 53 : 133—148. Dahlberg G. 1942. Methodik zur Unterscheidung von Erbeichkeits- und Milieuvariationen mit Hilfe von Zweillingen. Hereditas, 28 : 409—428. Dahlberg G. 1943/44. Genotypic asymmetries. Proc. Roy. Soc. Edinburgh, Sec. B, 62 : 20—31. Dahlberg G. 1948. Enwironment, inheritance and random variations with special reference to investigations of twins. Acta Genet. et Stat. Med., I : 104 —114. Dahlberg G. 1951. An explanation of twins. Sci. Amer., 184 : 48 — 51. Dahlberg G. I 952. Die Tendenz zu Zwillingsgeburten. Acta Gen. Med. Gemellolog., 1 : 80—87. Dareste C. 1891. Recherches stir la production artificielle des monstruosites. 2-me ed., Paris. Das K. 1934. Twin pregnancy, a demographic and ethnic study. J. Obst, Gynecol. Brit. Emp., 41 : 227—255. Davenport Ch. B. 1919. A strain producing multiple births. J. Hered., 10 : 382—384. Davenpоrt C. B. 1928. Is the inheritance of twinning tendency from the fathers side? Ztschr. ind. Abstamm.- u. Vererbungsl., 46 : 85—86. Davenport Ch. 1939. The genetical basis of resemblance in the form of the nose. Kultur u. Rasse : 60—64. Davis H. a. P. Davis. 1936. Action of the brain in normal persons and in normal states of cerebral activity. Arch. Neurol, a. Psychiatr., 36 : 1214. Day E. J. 1932. The development of language in twins. Child Developm., 179—199; 298—316. Detwiler S. 1933. Twinning in seven generations of the Bertolet family. J. Hered., 24 : 139 —142. Diehl К. u. O. Verschuer. 1933. Zwillingstuberkulose. Jena. Diehl К u. О. Verschuer. 1936. Der Erbeinfluss bei der Tubeikulose (Zwillingstuberkulose. II). Jena. Diesel P. 1954. Rh-Farktor und letale Erythroblastose bei diskordanten eineiigen Zwillingen. Dtsch. med. Wochenschr., 7, 31/32 : 958. Donald H. et al. 1951. Monozygotic bovine quadruplets. Heredity, 5 : 135—142. Driesch H. 1891. Entwicklungsmechanische Studien. I—II. Ztschr. wiss. Zool., 53 : 160—184. Driesch H. 1892. Entwicklungsmechanische Studien. III—IV. Ztschr. wiss. Zool., 54 : 1—62. Duis B. 1956. Zwillings Katamnesen. Acta Gen. Med. Gemellolog., 5 : 14—103. Duкe K. L. 1946. Monozygotic twins in the dog. Anat. Rec., 94 : 35 — 39. Duncan M. 1865. On some laws of the production of twins. Edinb. Med. Journ., 10 : 767. Dunsford J., С. Bоwleу, A. Hutchisоn, J. Thоmpsоn, R. Sanger a. R. Race. 1953. A human blood-group chimera. Brit. Med. Journ., 4827 : 81. Durr S. P. 1952. Craniopagus twins. Report of a case. J. Amer. Med. Ass., 150 : 93—94. Eitrem S. 1902. Die gottlicben Zwillinge bei den Griechen. Christiania. Enders R. a. O. Pearson. 1943. The blastocyte of the fisher. Anat. Rec., 85 : 285 — 287. Enders T. a. Stern. 1948. The frequencies of twins, relative to age of mothers, in American populations. Genetics, 33 : 263—272. Ernst A. 1901. Beitrage zur Kenntnis der Entwicklung des Embryosacks und des Embryo (Polyembryonie) von Tulipa gesneriana. Flora, 88 : 37 — 77. Essen-Moller E. 1941. Empirische Aehnlichkeitsdiagnose bei Zwillingen. Hereditas, 27 : 1—50. Eugster J. 1936. Zur Erblichkeitsfrage des endemischen Kropfes. III Teil. Die Zwillingsstruma. Arch. J. Klaus-Stift., 11 : 369—539. Eуster H. a. D. Вurpee. 1936. Inheritance of doubleness in the flowers of the Nasturtium. J. Hered., 27 : 51—60. Fernandez M. 1913. Die Entstebung der Einzelembryonen aus dem einheitlicben Keim beim Gurteltier Tatusia hybrida. IX Congr. Intern. Zool. Monaco : 401—414. Fierz F. 1952. Primartuberkulose bei eineiigen Zwillingen. Schweiz. Ztschr. Tuberk., 9 : 42—47. Fisher E. 1931. Die Entwicklungsgeschichle des Dachses und die Frage der Zwillingsbildung. Anat. Anz., 72, Verh. Anat. Ges. : 22— 34. Fischer H. 1956. Untersuchungen an Zwillingen von Beta vulgaris L. Zuchter, 26 : 136—152. Fischer R. G. 1952. Spina bifida and cranium bifidum. Forbes G. 1938. A case of thoracopagus monosymmetros. Anat. Rec., 70 : 237. Ford N. a. A. Mason. 1941. Taste reactions of the Dionne quintuplets. J. Hered., 32 : 365— 368. Ford N. a. A. Mason. 1943 Heredity as an etiological factor in malocclusion. As shawn by a study of the Dionne quintuplets. J. Hered., 34 : 57—64. Fougeroux. 1759. Un monstre forme de deux lapins reunis depuis la tete jusqu’au bas du sternum. Mem. Acad. Roy. Sci., Paris. Franceschetti A. et al. 1949. Angiomes tubereux a localisations diverses chez deux paires de jumeaux univitellins. Arch. J. Klaus Stift., 24 : 365—372. Frass F. 1932. Partial twinning in a hereford calf. J. Hered., 23: 88. Freeksen E. 1938. Die Venen des menschlichen Handruckens. Ztschr. Anat. u. Entwicklungsgesch., 108 : 91—99. Frischeisen-Kohler I. 1930. Untersuchungen an Schulzeugnissen von Zwillingen. Ztschr. f. angew. Psychol., 37 : 385—416. Galtоn Fr. 1875. The history of twins as a criterion of the relative powers of nature and nurture. J. Anthrop. Inst., 5 : 391—400. Galtоn Fr. 1895. Finger-print directories. London. Gardner I. a. H. Newman. 1940a. Mental and physical traits of identical twins reared apart. Case XX. Twins Lois and Louise. J. Hered., 31 : 119—126. Gardner I. a. H. Newman. 1940b. The alphabetical Perricone quadruplets. A Four-egg set, all males. J. Hered., 31 : 307— 314. Gardner I. a. H. Newman. 1940c. Physical and mental traits of the College quadruplets. J. Hered., 31 : 419—424. Gardner I. a. H. Newman. 1942. Studies of quadruplets. III. Types and frequencies of quadruplets. J. Hered., 33 : 9. Gardner I. a. H. Newman. 1943. Studies of quadruplets. VI. The only living one-egg quadruplets (Morlok). J. Hered., 34 : 258— 263. Gardner I. a. H. Newman. 1944. Studies of quadruplets. VII. The Schenses, four-egg quadruplets. J. Hered., 35 : 83—88. Gardner I. a. Rife. 1941. The diagnosis of five sets of triplets. J. Hered., 32 : 27—32. Gardner E. I., G. Stephens. 1949. Schizophrenia in monozigotic twins. J. Hered., 40 : 165—167. Gates R. R. 1946. Human Genetics, v. II. Ch. XXI Twins and twinning : 890—954. Gedda L. 1951. Studio dei gemelli. Roma : I—XVI+1—1384. Gedda L. 1956. Contributo alla storia dello studio dei gemelli. Acta Gen. Med. Gemellolog., 5 : 1—13. Gedda L. 1957. Il «metodo clinico-gemellare» nel rilievo del sintomo radiologico della stasi cerebrale e dell’influsso negativo delle cardiopatie sullo sviluppo corporeo (3 coppie MZ cardiopatiche discordante). Acta Gen. Med. Gemellolog., VI, 2 : 121 —146. Gedda L. et al. 1955. La voce dei gemelli. I. Prova di identificazione intragemonale della voce in 104 coppie (58 MZ e 46 DZ). Acta Gen. Med Gemellolog., 4 : 121—129. Gedda L. eNeroni. 1955. Reazioni posturali e mimiche di 56 coppie di gemelli. . . Acta Gen. Med. Gemellolog., 4 : 15—31. Geipel G. 1942. Die Verteilung der Fingerleistenmuster und die homologe Konkordanz bei ein- und zweieiigen Zwillingen. Ztschr. Morph, u. Anthrop., 40 : 51—79. Gemmill J. F. 1912. The teratology of fisches. Glasgow. Geoffroy Saint-Hilaire Is. 1837. Histoire generale et particuliere des anomalies de l’organisation chez l’homme et les animaxu etc. ou traite de teratologie. T. III. Bruxelles. Gernez L. et I. Оmez. 1938. L’accouchement gemellaire considers surtout du point de vue du pronostic et de la conduite a tenir. Rev. de gynec. et d’obst. : 916—941. Gesell A. 1921. Hemi-hypertrophy and mental defect. Arch. Neurol, a. Psychiatr. : 305—344. Gesell A. 1927. Hemi-hypertrophy and twinning. A futher study of the nature of hemi-hypertrophy with report of a case. Amer. J. Med. Sci., 173 : 542—555. Gesell A. 1942. The method of co-twin control. Science, 95 : 446—448. Gesell A. a E. Blake. 1936. Twinning and ocular pathology: with a report of bilateral macular colomba in monozigotic twins. Arch. Ophthalm., 15 : 1050—1070. Gesell A. a. H. Thompson. 1941. Twins T and C from infancy to adolescence: a biogenetic study of individual differences by the method of co-twin control. Genet. Psychol. Monogr., 24 : 3 —121. Gesell A. a. H. Thompson. 1952. Growth patterns of identical twins. Psychol. studies of human development, N. Y. : 16—24. Geyer H. 1931. Der Trinkversuch bei eineiigen und zweieiigen Zwillingen. Klin. Wochenschr., 10: 1488—1492. Geyer H. 1937. Uber den Schlaf von Zwillingen. Ztschr. ind. Abstamm.- u. Vererbungsl, 73, 3/4 : 524—527. Geyer H. 1939. Uber Hirnwindungen bei Zwillingen. Ztschr. Morph, u. Anthrop., 38 : 51—55. Gianferrari Luisa. 1954. Longevita ed eredita. Genetica medica. I symposium intern, gen. med. Roma : 19—40. Gigas H. 1941. Untersuchungen uber Muskelvarietaten an Zwillingen. Ztschr. Morph, u. Anthrop., 39 : 480—537. Glatzel H. 1931. Beitrage zur Zwillingspathologie. Ztschr. klin. Med., 116 : 632—668. Glatzel H. 1932. Untersuchungen von Magensekretion und Blutbild bei gesunden Zwillingen. Ztschr. ind. Abstamm.-u. Vererbungsl, 62 : Goetsch W. u. O. Schindler. 1934. Entwicklungsphysiologische Untersuchungen an Fischlarven. I. Beobachtungen und Versuche an Forellenzwillingen. Roux’ Arch., 131 : 483—511. Gottschaldt K. 1939. Erbe und Umwelt in der Entwicklung der geistigen Personlichkeit. Ztschr. Morph, u. Anthrop., 38 : 10—17. Gottschick J. 1939. Sprachpsychologische Zwillingsuntersuchungen. Arch. Ges. Psychol., 103 : 1—70. Graper L. 1931. Primitiventwicklung und einheitliche Erklarung von Doppelbildungen. Verb. anat. Ges., 40, Beih. anat. Anz., 72. Graewe H. 1950. Zwillingsforschung und Erbwissenschaft. Naturwissensch. Rundschau, 3. Graf L. 1939. Das Elektrokardiogramm bei ein- und zweieiigen Zwillingen. Ztschr. Kreislaufforsch., 31 : 337—349. Grebe H. 1952. Diskordanzursachen bei erbgleichen Zwillingen. Acta Gen. Med. Gemellolog., 1 : 89—102. Grebe H. 1955. Sport bei Zwillingen. Acta Gen. Med. Gemellolog., 4 : 275—294. Grebe H. 1956. Fortschritte in der Erforschung mutterlicher Krankheiten als Ursache kindlicher Missbildungen. Naturwissensch. Rundschau, 9 : 49—52. Green С. V. 1934. Occurence of uniovular twins in multiple births. Science, 80 : 616. Greene J. 1942. Atypical laryngeal and vocal changes in adolescence. J. Amer. Med. Ass., 120 : 15. Greulich W. W. 1930. The incidence of human multiple births. Amer. Nat., 64 : 142 —153. Greulich W. W. 1934. Heredity in human twinning. Amer J. Phys. Anthrop., 19 : 391—431. Greulich W. W. 1938. The birth of six pairs of fraternal twins to the same parents. J. Amer. Med. Ass., 110, 8 : 559—563. Grimm J. u. W. 1850. Die zwei Bruder. Kinder- und Hausmarchen. Bd. I. Gottingen : 360. Grosser О. u. H. Przibram. 1906. Einige Missbildungen beim Dornhai (Acanthias vulgaris Russo). Arch. Entw.-Mech., 22 : 21—37. Grunwald P. 1942. Early human twins with peculiar relations to each other and the chorion. Anat. Rec., 83 : 267—269. Guldberg E. 1938. Verschiedengeschlechtige eineiige Zwillinge. Acta path. scandinav., Suppl., 37 : 197—223. Guttmаскer A. 1948. Type-knapping and the Ohio octuplets. J. Hered., 39 : 294. Hаак D. 1943. Untersuchungen an eineiigen und zweieiigen Rinderzwillingen. Ztschr. Tierzuchtung u. Zuchtungsbiol., 54 : 27—77. Habs H. 1937. Zur Erbbedingheit der Dunkeladaption. Ztschr. ind. Abstamm.-u. Vererbungsl., 73, 3/4 : 531. Habs H. 1938. Zwillingsphysiologische Untersuchungen uber die Erbbedingtheit der alveolaren CO2-spannung u s. w. Ztschr. menschl. Vererb.-u. Konstitutionsl., 21 : 447—475. Habs H. 1939. Krebs und Vererbung. Ztschr. f. klin. Med., 135 : 679—691. Habs H. 1948. Massenstatistik und Zwillingsmethode in ihrem Wert fur die Analyse biologischer Probleme. Grenzgeb. d. Mediz., 1 : 106—108. Haccius B. 1955. Experimentally induced twinning in plants. Nature, 176, 4477 : 355—356. Hall H. 1928. Twins in Upper Guinea. Museum J. Univ. Pensylv. Hamlett G. 1935a. Human twinning in the Un. States: racial frequencies, sex ratios and geographical variations. Genetics, 20 : 250—258. Hamlett G. 1935b. Delayed implantation and discontinuous development in the mammals. Quart. Rev. Biol., 10 : 432—447. Hamlett G. a. G. Wislocki 1934. A proposed classification for types of twins in mammals. Anat. Rec., 61 : 81—96. Hammоnd J. 1941. Fertility in mammals and birds. Biol. Reviews, 16 : 165—190. Hammоnd J. 1949. Induced twin ovulations and multiple pregnancy in cattle. J. Agric. Sci., 39 : 222—225. Hanсоск J. 1954. Monozygotic twins in cattle. Advanc. in Genetics, VI: N. Y. : 141—181. Hanhart E. 1953. Eineiige Zwillingsmadchen mit konkordantem Albinismus universalis. Acta Gen. Med. Gemellolog., 2 : 380—390. Hansson A. 1947. The physiology of reproduction in mink (Mustela vison, Schreb.) with special reference to delayed implantation. Acta Zool.. 28, 1. Harland S. 1936. Haploids in polyembryonic seeds of Sea island cotton. J. Hered., 27 : 229—231. Harris R. 1892. The blended Tocci brothers and their historical analogues. Amer. J. Obst. Diseas, of Women a. Childr. (N. Y.), 25, 171 : 460—473. Harris R. 1957. Refractive errors in monosygotic twins. Eye, ear, nose and throat monthly, 36 : 41—42. Harris M., J. Rendel. 1903. The Dioscuri in the Christian legends. London. Harris M., J. Rendel. 1906. The cult of the heavenly twins. Cambridge. Hartmаn C. G. 1920. The free-martin and its reciprocal: opossum, man, dog. Science, 52 : 469—471. Hartmаn C. G. 1943. Birth of a two-headed monster in the Rhesus monkey. Science, 98 : 449. Harvey E. U. 1940. A new method of producing twins, triplets and quadriplets in Arbacia punctulata and their development. Biol. Bull., 78 : 2. Harvey G. 1651. Exercitationes de generatione animalium. Amsterdam. Hauenschild C. 1954. Genetische und entwicklungsphysiologische Untersuchungen uber Intersexualitat und Gewebevertraglichkeit bei Hydractinia echinata. Roux’Arch., 147 : 1—41. Heaney N. а. Вartelmez. 1931. A case of monochorial twins with a single yolk sac. Anat. Rec., 48, Suppl. : 47. Hellin D. 1895. Die Ursache der Multiparitat der uniparen Tiere uberhaupt und der Zwillingschwangerschalt beim Menschen. Munchen. HeIweg-Larsen H. 1947. Situs inversus in one monozigotic twin. Ann. Eugen., 14 : 1—8. Henning W. 1937. A double sheep pregnancy with a single corpus luteum. J. Hered., 28 : 61—62. Herbst C. 1900. Uber das Auseinandergehen von Furchungs- und Gewebszellen in kalkfreiem Medium. Arch. Entw.-Mech., 9 : 424—463. Hermann E. 1940. Messungen an Handschriftproben von Zwillingen unter 14 Jahren. Ztschr. Psychol., 147 : 238—255. Herrman L. 1953. Finger-prints and twins, with special reference to identical twins. J. Forensic Med., 1 : 101. Hertwig O. 1893. Uber den Wert der ersten Furchungszellen fur die Organbildung des Embrio. Arch. mikr. Anat., 42 : 662—806. Hess H. 1938. Beitrage zur Kenntnis der Asymmetrie des Gesichts bei menschlichen Embryonen, Neugeborenen und bei Kindern bis zum 14 Lebensjahr. Ztschr. Morph. u. Anthrop., 37 : 567. Hey A. 1911. Uber kunstlich erzeugte Janusbildungen von Triton taeniatus. Arch. Entw.-Mech., 33: 117—195. Hildebrand S. 1938. Twinning in Turtles. J. Hered., 29 : 243—253. Hilgard J. 1933. The effect of early and delayed practice on memory and performances studied by the method of co-twin control. Genet. Psychol. Monogr., 14 : 493—565. Hinrichs M. 1938. The microscopic anatomy of twins and double monsters of Fundulus heteroclitus. Physiol. Zool., 11 : 155—157. Hinrichs M. a. J. Genther. 1931. Ultraviolet radiation and the production of twins and double monster. Physiol. Zool., 4 : 461—485. Hirst J. C. 1940 Problem of multiple pregnancy. Pennsylv. Med. Journ., 43 : 1553. Hirt L. 1902. Zur Kenntnis der Zwillingsschwangerschaft. Diss., Breslau. HоadIeу L. 1928. Twin heterosexual pig embryos formed with fused membranes. Anal. Rec., 38 : 177—187. Hollander R. 1954. Die monoamniotische Zwillingsschwangerschaft. Zbl. Gynak., 74 : 409—414. Hollander R. u. Drescher. 1954. Uber die Beziehungen zwischen Schwangerschaftsdauer und Reifezeichen bei Zwillingen. Ztschr, Geburtshilfe, u. Gynak, 140 : 55—65. Ноlstе А. 1940. Die Refraktionsverhaltnisse bei ein- und zweieiigen Zwillingen. Arch. Ophthalm., 142 : 467—473. Home E. 1790. An account of a child with a double head. Philos. Trans. Roy. Soc. London, 80 : 296—305. Hormont C. 1954. Research with cattle twins. In: Statistics and mathematics in biology. Jowa, USA : 407—418. Horstadius S. 1928. Uber die Determination des Keimes bei Echinodermen. Acta Zool., 9 : 1. Huber A. 1931. Ophthalmologisch Untersuchungen an ein- und zweieiigen Zwillingen. Arch. J. Klaus-Stift., 6 : 41—122. Hubner H. 1911. Die Doppelbildungen des Menschen und der Tiere. Ergebn. d. allgem. Pathol, u. patholog. Anat. d. Menschen u. d. Tiere, 15 Jahrg., II Abt. : 650—796. Hutt F. B. 1930. Bovine quadruplets. J. Hered., 22 : 339—348. Hutter K. 1953. Folgen verschiedener Handigkeit bei erbgleichen Zwillingen. Wien. klin. Wochenschr., 12 : 232. Jablonsky W. 1922. Ein Beitrag zur Vererbung der Refraktion menschlicher Augen. Arch. f. Augenheilk., 91 : 308. Janскe G. 1941. Die Augenmerkmale bei Zwillingen und ihre Bedeutung fur die Diagnose der Ein- der Zweieiigkeit. Graef. Arch. Ophthalm., 142 : 113—155. Jarviк L. et al. 1957. Changing intellectual functions in senescent twins. Acta genet. et statist, med., 7 : 421—430. Jenkins R. a. J. Gwin. 1940. Twins and triplet birth ratios J. Hered., 31 : 243—248. Jennings D. 1937. Identical twins. Lancet, 233 : 655. Jespersen O. 1925. Die Sprache, ihre Natur, Entwicklung und Entstehung. Heidelberg. Jeune M. et J. Confavreux. 1948. Une paire de jumeaux monoamniotiques en miroir. (Sit. inv. et maladie bleue chez l’un d’eux). Arch. franc. de pediatr., 5 : 252—256. Johansen D. A. 1951. Plant empbryology (Embryol. of the Spermatophyta). Mass. USA. Johansson I. 1932. The sex ratio and multiple births in cattle. Ztschr. f. Zuchtg., 24 : 183—268. Johansson J. a. O. Venge. 1951. Studies on the value of various morphological characters for the diagnosis of monozygocity of cattle twins. Ztschr. Tierzucht. u. Zuchtungsbiol., 59 : 389—424. Johnson D. 1944. Quintuple lambs of woolless domestic sheep. J. Hered., 35 : 251—253. Jones H. a. D. Morgan. 1942. Twin similarities in eye movement patterns. J. Hered., 33 : 167—172. Кadanoff D. 1940. Untersuchungen uber die Unterschiede in der Verastelung der Hautnerven und Venen der oboren Extremitat bei ein- und zweieiigen Zwillingen. Ztschr. Morph, u. Anthrop., 38 : 73—89. Kaestner S. 1907. Doppelbildungen an Vogelkeimscheiben. Arch. Anat. u. Entwicklungsgesch. : 129—215. Kahler O. u. Weber. 1940. Zur Erbpathologie von Herz- und Kreislauferkrankungen. Ztschr. klin. Med., 137, I : 380—417; II : 507 575. Kallmann Fr. 1950. The genetics of psychoses. An analysis of 1, 232 twin index families. Amer. J. Hum. Gen., 2 : 385—390. Kallmann Fr. 1952a. Comparative twin study of the genetic aspects of male homosexuality. J. Nerv. a. Ment. Disease, 115 : 283—298. Kallmann Fr. 1952b. The genetic aspects of mental disorders in the aging. Comparative twin data on the involutional and senile periods of life. J. Hered., 43 : 89—96. Kallmann F. 1953. Heredity in health and menial disorder. N. Y. Kallmann F. u. D. Reisner. 1943. Twin studies on genetic variations in resistance to tuberculosis. J. Hered.: 269—276. Kallmann F. a. G. Sander. 1948. Twin studies on aging and longevity J. Hered., 39 : 349—357. Kallmann F., B. Aschner a. A. Falik. 1956. Comparative data on longevity, adjustment to aging and causes of death in a senescent twin population. Novant’Anni delle leggi mendeliane. Roma : 330—339. Кammerer Th. 1950. Trois cas d’epilipsie concordante chez des jumeaux. L’encephale, 39, 6 : 568—579. Kappert H. 1933. Erbliche Polyembryonie bei Linum usitatissimum. Biol. Zbl., 53 : 276—307. Kappert H. 1950. Botanische Untersuchungen zur Erblichkeit der Polyembryonie. Mod. Biol. (Festschr. Nachtsheim). Kean В. H. 1942. Complete transposition of the viscera in both of oneegg twins. J. Hered., 33 : 217—221. Keeler С. E. 1929. On the amount of external mirror imagery in double monsters and identical twins. Proc. nat. Acad. Sci. USA, 15 : 839—842. Keen J. A. 1947. Anatomy and embryology of double monsters, with special reference to dicephalus. S. Afr. J. med. Sci., 12 : 117— 139. Keiter F. 1950. Uber Zehenbeerenmuster und Kompliziertheitsindex. Ztschr. Morph, u. Anthrop., 42 : 169—203. Keller K. u. Th. Niedоba. 1937. Untersuchungen an Doppelmonstren des Rindes im Sinne der Zwillingsforschung. Ztschr. Zuchtung, (Reibe B), 37 : 245—293. Кiil V. 1948. Inheritance of frontal hair direction in man. J. Hered., 39 : 206—216. Kimball О. a. Hersh. 1955. The genetics of epilepsy. Acta Gen. Med. Gemellolog., 4 : 131—141. King J. W. 1953. A feeding experiment with twin cattle. Proc. Brit. Soc. An. Prod., : 76. Kleinenberg N. 1879. The development of the earthworm Lumbriculus trapezoides. Quart. J. Microsc. Sci., 19 : 206—244. Kleinwachter L. 1871. Die Lehre von den Zwillingen. Prag. Knoch J. 1873. Ueber Missbildungen betreffend der Embryonen des Salmonen- und Goregonus-Geschlechts. Bull. Soc. des Naturalistes de Moscou, XLVI : 2. Коmai T. 1928. Criteria for distinguishing identical and fraternal twins. Quart. Rev. Biol., 3 : 408—418. Коmai T. 1938. Problem of situs inversus viscerum, as studied on single and duplicate salmon embryos. Mem. Coll. Sci. Kyoto Imp. Univ., ser. B., 14 : 155—170. Коmai T. a. G. Fuкuокa. 1934a. A note on the problem of mirrorimaging in human twins. Hum. Biol., 6 : 24—32. Коmai T. a. G. Fuкuокa. 1934b. A. study of the frequency of left-handedness and left-footedness among Japanese school children. Hum. Biol., 6 : 33—42. Komai T. a. G. Fukuoka. 1934c. Post-natal growth disparity in monozygotic twins. J. Hered., 25 : 423—430. Komai T. a. G. Fukuoka. 1936. Frequency of multiple births among the Japanese and related peoples. Amer. J. Phys. Anthrop. 21 : 433—447. Коpsch Fr. 1897. Uber eine Doppelgastrula bei Lacerta agilis. Sitzungsber. Akad. Wiss. Berlin, 29 : 1—5. Коrкhaus G. 1929a. Die erste Dentition und der Zahnwechsel im Lichte der Zwillingsforschung. Vierteljschr. Zahnheitk., 45 : 414— 430. Коrкhaus G. 1929b. Die Vererbung der Anomalien der Zahnzahl. Korresp.-bl. Zahnarzte, 11 : 1—24. Коrкhaus G. 1930. Die Vererbung der Zahnfarbe. Ztschr. Ges. Anat. u. Konstitutionsl., 15 : 127. Коrкhaus G. 1931. Aetiologie der Zahnstellungs und Kieferanomalien. Fortschr. Orthod., I. Коrкhaus G. 1942. Uber den Einfluss der Erbmasse auf das Gebiss. (Befunde an Drillingen und Vierlingen). Verb. d. 23. Kongr. d. Europ. Orthod. Gesel., Leipzig. Коrкhaus G. 1954. Genotypische und paratypische Einflusse beim Aufbau von Gebiss und Gesichtsschaedel. Genetica medica. Red. L. Gedda. Roma : 344—351. Korschelt E. 1927. Regeneration und Transplantation Bd. I. Regeneration. Berlin : 414 Krause G. 1952. Schnittoperation im Insektenei zum Nachweis komplementarer Induktion bei Zwillingsbildung. Naturwissenschaften, 39 : 356. Krause G. 1953. Die Aktionsfolge zur Gestaltung des Keimstreifs von Tachycines (Saltoria), insbesondere das morphogenetische Konstruktionsbild bei Duplicitas parallela. Roux’Arch., 146 : 3. Kronacher G. 1932. Zwillingsforschung bei den Haustieren, im besonderen beim Rind. Ztschr. ind, Abstamm. u. Vererbungsl., 62 : 126—152. Kronacher C. u. F. Hоgreve. 1936. Rontgenologische Skeletstudien an Dahlemer Rinder—Drillingen und Zwillingen. Ztschr. Zucht., 36 : 281—294. Kronacher G. u. D. Sanders. 1936. Neue Ergebnisse der Zwillingsforschung beim Rind. Ztschr. Zucht., 34 : 1—83, 133—220. Кuhne K. 1936. Die Zwillingswirbelsaule. Ztschr. Morph, u. Anthrop., 35 : 1—376. Lasкer G. а. Reуnоlds. 1948. A cicatrix of the neck associated with anomalous dental occlusion in one of a pair of monozygotic twins. Hum. Biol., 20 : 36—46. Lamy M. et J. Frezal. 1959. Etudes sur l’etiologie de la gemellite chez l’homme. X intern. Congress of genet, vol. I : 89—99. Lassen M. T. 1931. Nachgeburtsbefunde bei Zwillingen und Aenlichkeitsdiagnose. II. Arch. Gynakol., 147 : 48—64. Lebegue A. 1952. La polyembryonie chez les angiospermes. Bull. Soc. Bot. de France, 99 : 329—367. Lee W. 1956. A study of the inheritance of certain tongue characters. J. Hered., 47 : 17—19. Lehmann W. 1938. Zwillings- und Familien-Untersuchungen zur Erbpathologie der Hyperthyreose. Ztschr. menschl. Vererb.-u. Konstitutionsl., 22 : 183—223. Lehmann W. 1954. Erbpathologie der Krankheiten der inneren Secretion. Genetica medica, Roma : 79—97. Lehmann W. u. J. Hartlieb. 1937. Capillaren bei Zwillingen. Ztschr. menschl. Vererb.-u. Konstitutionsl., 21 : 21. Lehmann F. u. Huber. 1944. Beobachtungen an Tubifex uber die Bildung von Doppeleiern bei der zweiten Reifungsteilung und die Frage der Entstehung ovozytarer Zwillinge. Arch. J. Klaus-Stift., 19 : 473. Lehmann W. u. Witteler. 1935. Zwillingsbeobachtungen zur Erbpathologie der Polydaktylie. Zbl. Chir., 62 : 2844—2852. Lehrfield L. 1948. Homologous twins foretold by eye grounds. Eye, Ear a. Mouth, 27 : 319. Leicher H. 1929. Uber die Vererbung der Nasenform. Verh. Ges. phys. Anthrop., 3. Lemery N. 1740. Troisieme memoire sur les monstres a deux tetes etc. Hist. Acad. Roy. Sci. Paris : 607—612. Lennоx W. G. 1951. The heredity of epilepsy as told by relatives and twins. J. Amer. Med. Ass., 146 : 529—536. Lennоx W. G. 1956. Transmission of the cerebral dysrhythmias and seizures of epilepsy. Novant’anni delle leggi mendeliane, Roma : 303—310. Lennоx W. a. A. Collins. 1945. Intelligence of normal and epileptic twins. Amer. J. Psych., 101 : 764—769. Lennоx W. G. a. F. Gibbs. 1945. The brain-wave pattern, an hereditary trait. J. Hered., 36 : 233—243. Lennоx W. a. Jolly. 1954. Seizures, brain waves and intelligence tests of epileptic twins. In: Genet. and the Inher. of Integrated Neurolog. and Psychiatric Patterns, Res. Publ., A. Nerv. a. Ment. Dis. 33 : 325. Lenz Fr. 1933. Zur genetischen Deutung von Zwillingsbefunden. Ztschr. ind. Abstamm.- u. Vererbungsl., 52. Lenz F. u. O. Verschuer. 1928. Zur Bestimmung des Anteils von Erbanlage und Umwelt an der Variabilitat. Arch. f. Rassen-u. Ges.-Biol., 20 : 425. Letovaara A. 1938. Psychologische Zwillingsuntersuchungen. Ann. Acad. Sci. Fenn. (Helsinki), Ser. B, 39 : 460. Levine Ph. 1954. The genetics of the newer human blood factors. Advan. in Genet., 6 : 183—234. Lezine I. 1953. Note sur le developpement psychomoteur de jeunes jumeaux. Sem. hopit., 29 : 3967—3973. Licetus F. 1665. De Monstris. Editio novissima, iconibus illustrata. Amsterodami. A. Frisii : 1—316. Liebenam L. 1938. Zwillingspathologische Untersuchungen aus dem Gebiet der Anomalien der Korperform. Ztschr. Konstitutionsl., 22 : 373. Lillie F. R. 1917. The free-martin, a study of the action of sex hormones in the foetal life of cattle. J. Exper. Zool., 23 : 371. Lillie F. R. 1923. Supplementary notes on twins in cattle. Biol. Bull., 44 : 47—78. LindahI P. E. 1937. Uber eineiige Zwillinge aus Doppeleiern. Biol. Zbl., 57 : 389—393. Little S. a. N. Weaver. 1950. Epilepsy in twins. Amer. J. Dis. Child., 79 : 223—232. Loeb J. 1894. Uber eine einfache Methode, zwei oder mehr zusammengewachsene Embryonen aus einem Ei hervorzubringen. Pflug. Arch., 55 : 525—529. Loeb J. 1909. Uber die chemischen Bedingungen fur die Entstehung eineiiger Zwillinge beim Seeigel. Arch. Entw. Mech., 27 : 119—140. Loewy E. 1925. Beobachtungen an Zwillingen: gibt es eineiige Zwillinge mit verschiedener Kopfhaarfarbe? Med. Klin., Berlin, 21 : 1125 Long J., О. Gilmore a. C. Rife. 1956. Heritability of thyroid status as indicated by the protein-bound iodine in the blood serum of man and cattle. Novant’anni delle leggi mendeliane Roma : 188—191. Lottig H. 1931. Hamburger Zwillingsstudien. Beih. Ztschr. angew. Psychol. Leipzig. Lotze R. 1937. Zwillinge. Einfuhrung in die Zwillingsforschung. Oehringen : 1—176. Lowe G. a. Th. Mс Кeоwn. 1954. An investigation of dextrocardie with and without transposition of abdominal viscera, with a report of a case in one monozygotic twin. Ann. Eugen., 18 : 267—277. Lowenstein 0. 1939. Les troubles du reflexe pupillaire a la lumiere dans les affections syphilitiques du systeme nerveux central. Paris. Luchsinger R. 1940. Die Sprache und Stimme von ein- und zweieiigen Zwillingen in Beziehung zur Motorik und zum Erbcharakter. Arch. J. Klaus-Stift., 15 : 461—527. Luchsinger R. 1944. Erbbiologische Untersuchungen an ein- und zweieiigen Zwillingen in Beziehung zur Grosse und Form des Kehlkopfs. Arch. J. Klaus-Stift., 19 : 393—441. Luchsinger R. 1953. Die Sprachentwicklung von ein- und zweieiigen Zwillingen und die Vererbung von Sprachstorungen. Acta Gen. Med. Gemellolog., 2 : 31—47. Ludwig W. 1932. Das Rechts-Links-Problem im Tierreich und beim Menschen. Berlin : 1—496. Ludwig W. 1949. Symmetrieforschung im Tierreich. Stud. Gen., 2 : 231—239. Lund S. 1933a. A psycho-biological study of a set of identical girl-triplets. Hum. Biol., 5 : 1—34. Lund S. 1933b. Determining similarity of triplets and quadruplets. J. Hered., 24 : 195—197. Lundstrom A. 1948. Tooth size and occlusion in twins. Stockholm. Luscher E. 1944. Otomikroskopische Beobachtungen an Zwillings-troinmelfellen. Arch. J. Klaus-Stift., 19, 461—462. Lush R. 1925. Inheritance of twinning. J. Hered., 16 : 273—279. Luth K. F. 1937. Endokrine Storungen bei eineiigen Zwillingen. Zlschr. Konstitutionsl., 21 : 55. Luther W. 1935. Entwicklungsphysiologische Untersuchungen am Forellenkeim. Die Rolle des Organisationszentrums bei der Entstehung der Embryonalanlage. Biol. Zbl., 55, 3/4. Lutz H. 1953. L’orientation des axes embryonnaires dans la gemellite experimentale chez les oiseaux et son determinisme. Bull. Biol., 87, 1. Lutz H. et Lutz-Ostertag. 1952. Sur la production experimentale de jumeaux chez l’embryon d’un Salmonide. C. R. Acad. Sci., 234 : 469—471. Luxenburger H. 1930. Psychiatrisch-neurologische Zwillingspathologie. Referat. Ztschr. ges. Neurol. u. Psychiatr., 56 : 145—180. Luxenburger H. 1935. Untersuchungen an schizophrenen Zwillingen und ihren Geschwistern zur Prufung der Realitat von Manifestationsschwankungen. Ztschr. ges. Neurol, u. Psychiatr., 54 : 351 — 394. Lynn W. G. 1946. Situs inversus viscerum in conjoined twins of the Brook Trout. J. Morph., 79: 1—25. Lуnn W. a. A. Peadоn. 1949. Situs inversus in conjoined triplets of the Brook Trout. J. Morph., 84 : 411—426. McArthur J. W. 1938a. Diagnosis of the Dionne quintupletsas a monozygotic set. J. Hered., 29 : 323—329. McArthur J. W. 1938b. Reliability of dermatologlyphics in twin diagnosis. Hum. Biol., 10 : 12—35. McArthur J. W. 1942. Relation of body size to litter size and to the incidence of fraternal twins. J. Hered., 33 : 87—91. McArthur N. 1954. Changes in twin frequencies. Acta Gen. Med. Gemellolog., 3 : 29—33. McArthur J. W. a. O. McArthur. 1937. Finger, palm and sole prints of monozygotic quadruplets. J. Hered., 28 : 147—153. McArthur J. W. a. A. Dafoe. 1939. Genetics of quintuplets. II. Trends of growth in the Dionne quintuplets. J. Hered., 30 : 359—364. Macfarlan D. 1927 Identical hearing in identical twins. Laryngoscope, 37 : 846. McFarland J. 1943. The problem of cancer in twins. Bull. Am. Soc. Control Cancer, 25 : 30—43. McKinney H. a. Sanda. 1934. Twisted wheat and twisted trees. Twisting of the rachis of wheat heads by short daily photoperiods suggest a new appoach to studies ot twistet trees. J. Hered., 25 : 261—263. Macklin M. 1934. A visit to Corbeil. J. Hered., 25 : 420—422. Macklin M. T. 1940. An analysis of tumors in monozygous and dizygous twins. J. Hered., 31 : 277—290. Mаскlin M. a. Mооr. 1935. An example of a similar type of malocclusion in identical twins. J. Hered., 26 : 444—450. McLaren D. W. 1936. Separation of conjoined twins. Brit. Med. Journ. : 971. Maheshwari P. 1950. An introduction to the embryology of angiosperms. N. Y. Maheshwari P. 1952. Polyembryony in angiosperms. Paleobotanist, 1 : 319. Mainland D. 1928/1929. Posterior duplicity in a dog, with reference to mammalian teratology in general. J. Anat. London, 63 : 473—495. Malan M. 1940. Zur Erblichkeit der Orientierungsfahigkeit im Raum. Ztschr. Morph, u. Anthrop., 39 : 1—23. Mangold O. 1929. Das Determinationsproblem. II. Ergebn. Biol., 5 : 290—404. Mangold О. u. I. Testa. 1953. Eineiige Zwillinge bei Triton, ihre experimentelle Herstellung und ihre Regulation. Acta Gen. Med. Gemellolog., 2 : 49—69. Mangold О., I. Testa, C. Woellwarth. 1956. Uber die Regulation der halbseitigen Organlagen bei kunstlich erzeugten Zwillingen von Triton alpestris. Acta Gen. Med. Gemellolog., 5 : 369—401. Marchand F. 1881. Die bohmischen Schwestern Rosalia und Josepha. Breslau aerztl. Ztschr., 3 : 237—240. Marchand F. 1897. Die Missbildungen. Eulenburgs Realenzykl. d. Ges. Heilkunde. Martin E. 1880. Histoiro des monstres depuis l’antiquite jusqu’a nos jours. Paris. Mattisson K. 1933. Zur Frage der Hereditat bei Situs inversus visserum totalis. Ztschr. Konstitutional., 17 : 325. Mayer-List R. u. G. Hubener. 1925. Die Capillarmikroskopie in ihrer Bedeutung zur Zwillingsforschung. Munch. med. Wochenschr., 72 : 2185. Meckel J. F. 1815. De duplicate monstrosa commentarius. Halle—Berlin. Melsоm R. 1944. Dermatological investigations on 22 pairs of identical twins. Acta dermat.-vener., 25 : 29—47. Meу R. 1950. Uteruskarzinom und Zwillingsforschung. Zbl. Gynakol., 72 : 1579—1583. Meyer H. 1932. Zur Vererbung der Zwillingsschwangerschaft. Arch. Rass.- u. Ges.-Biol., 26 : 387—417. Meуer-Heуdenhagen G. 1934a. Die Palmaren Hautleisten bei Zwillingen. Ztschr. Morph. u. Anthrop., 33 : 1—42. Meуer-Heуdenhagen G. 1934b. Zur Erbbiologie der Papillarleisten der Handflache. Ztschr. ind. Abstamm- u. Vererbungsl., 67 : 302—305. Miettinen M. 1954. On triplets and quadruplets in Finland. Acta Paed., 43, Suppl. 99 : 1 —103. Miller R. E. 1941. A study of quadruplets. Anat. Rec., 80 : 411—420. Mitschrich H. 1955. Zwillingstuberkulose. III. Nachuntersuchung nach 20 Jahren an der Serie tuberkuloser Zwillinge von K. Diehl und 0. Verschuer. Stuttgart. Mоndу G. 1953. Tuberkulose bei Zwillingen. Beitr. klin. Tuberk., 109 : 183 — 186. Morgan D. 1939. Twin similarities in photographic measures of eye movements while reading prose. J. Educ. Psychol., 30 : 527— 586. Morgan D. 1951. Polyembryony in maize and lily. J. Hered., 42 : 91—93. Morgan D. a. R. Rappleуe. 1950. Twin and triplet pepper seedlings. A study of polyembryony in Capsicum frutescens. J. Hered., 41 : 91—95. Morgan T. H. 1927. Experimental embryology. N. Y. : 317 сл. Mоriкawa N. 1939. Topographisch-anatomische Untersuchung uber den Magen der japanischen Zwillingsfoten. Okajimas Folia anat. japon., 18 : 163—197. Morita S. 1936. Die kunstliche Erzeugung von Einzelmissbildungen von Zwillingen, Drillingen und Mehrlingen im Huhnerei. Anat. Anz., 82 : 81—160. Morley M. 1957. The development and desorders of speech in Chilodhood. Edinburgh a. London : 1—440. Morrill C. N. 1919. Symmetry reversal and mirrorimaging in monstrous trout and a comparison with similar conditions in human double monsters. Anat. Rec., 16 : 4. Mоrsier G. 1945. Psychose hallucinatoire identique chez deux jumelles univitellines. Arch. J. Klaus-Stift., 19 : 468—472. Mundу G. 1953. Tuberkulose bei Zwillingen. Beitr. klin. Tuberk., 109 : 183. Nolbandov A. 1942. A case of viable twin chicks. J. Hered., 33 : 53— 54. Nehls G. 1940. Caries und Paradendentose bei Zwillingen. Ztschr. menschl. Vererb- u. Konstitutionsl., 24 : 235—247. Nelson S., N. Hunter a. M. Walter 1945. Stuttering in twins. J. Speech Disorders, 10 : 335—343. Newman H. 1923. The physiology of twinning. Chicago. Newman H. 1928a. Studies of human twins. I. Methods of diagnosing monozygotic and dizygotic twins. Biol. Bull., 55 : 283—297. Newman H. 1928b. Studies of human twins. II. Asymmetry reversal, or mirror-imaging in identical twins. Biol. Bull., 55 : 298—315. Newman H. 1930. The finger prints of twins. J. Genet., 23 : 415—446. Newman H. 1931. Palm-print patterns in twins. J. Hered., 22 : 40—49. Newman H. 1940a. Twin and triplet chick embryos. J. Hered., 31 : 371—378. Newman H. 1940b. The question of mirror imaging in human one-egg twins. Hum. Biol., 12 : 21—34. Newman H. 1940c. Multiple human births. N. Y. Newman H. 1948. Report of octuplets and sextuplets in the Un. States. J. Hered., 39 : 204—205. Newman H. a. J. Patterson. 1909. A case of normal identical quadruplets in the nine-banden Armadillo, and its bearing on the problems of identical twins and of sex determination. Biol. Bull., 17 : 3. Newman H. a. W. Quisenberry. 1944. One-egg twins with spina bifida and polydactyly. J. Hered., 35 : 309—314. Newman H., F. Freeman, К. Hоlzinger. 1937. Twins. A study of heredity and environment. Chicago : 369. Nichоlas J. a. Hall. 1942. Experiments on developing rats. II. The development of isolated blastomeres and fused eggs. J. exp. Zook, 90 : 441—458. Niсоlle C. 1935. A propos d’un cas de fecondite quadrigemellaire chez une femme de race pygmee. Bull. Soc. obscetr. gynecol., 24. Nixon B. 1956. On the diagnosis of twin-pair ovularity and the use of dermatoglyphic data. Navant’anni delle leggi mendeliane. Roma : 235—245. Oppenheimer J. 1936. Transplantation experiments on developing Teleosts (Fundulus and Perca). J. ex. Zool., 72 : 3. Orth H. 1954. Extreme Discordanz der Refraktionswerte eineiiger Zwillinge. Klin. Monatsbl. Augenheilk., 124 : 304—306. Оstertag M. 1936. Die Abhangigkeit des Erythrocytendurchmessers und des Blutbilds von erblichen Einflussen nach Untersuchungen an Zwillingen. Dtsch. Arch. klin. Med., 178 : 201—206. Ostlyngen E. 1949. Possibilities and limitations of twin research as a means of solving problems of heredity and environment. Acta Psychol., 6 : 59—90. Owen R. D. 1945. Immunogenetic consequences of vascular anastomosis between bovine twins. Science, 102 : 400. Owen R., H. Dawis a. R. Morgan. 1946. Quintuplet calves and erythrocyte mosaicism. J. Hered., 37 : 291—297. Panaghioton P. 1938. Eineiige Zwillinge verschiedener Blutgruppen. Dtsch. med. Wochenschr., 64 : 1544 — 1545. Panum P. L. 1860. Untersuchungen uber die Entstehung der Missbildungen zunachst in den Eiern der Vogel. Kiel. Parade G. u. W. Lehmann 1939. Elektrokardiogramme bei Zwillingen. Ztschr. menschl. Vererb., 22 : 96—104. Pare A. 1578. De la generation de l’homme. Les monstres tant terrestres que marins. Paris. Patten B. 1946. Human embryology. Philadelphia : 204—219. Patterson J. T. 1913. Polyembryonic development in Tatusia novemcincta. J. Morph., 24 : 559—662. Patterson J. T. 1927. Polyembryony in animals. Quart. Rev. Biol., II, 3 : 399—426. Patterson J. T. a. Hartman. 1917. A polyembryonic blastocyst in the opossum (Didelphys virginiana). Anal. Rec., 13. Patzig B. 1937. Zur Vorerbung der Schadelformen. Ztschr. ind. Abstamm.-u. Vererbungl., 73 : 550—559. Peiреr А. 1923. Zur Vererbung der Zwillingsschwangerschaft durch den Mann. Klin. Wochenschr., II : 1651. Penners A. 1924. Doppelbildungen bei Tubifex rivulorum. Zool. Jahrb., Abt. allg. Zool., 41 : 91 — 120. Perlstein M. a. E. Leсоunt. 1927. Pygopagus twins; history and necropsy report of bohemian twins Rosa-Josepha Blazek. Arch. Path. a. Lab. Med., 3 : 171. Petri E. 1934. Untersuchungen zur Erbbedingtheit der Menarche. Ztschr. Morph, u. Anthrop., 33 : 43—48. Peyron A. et N. Kobozieff. 1937. Les tumeurs cher les jumeaux. Bull. Assoc. franc. Cancer, 26 : 93—110. Pezzi C. e L. Carugati. 1924. Destrocardia e trasposizione viscerale (Situs inversus) in due geinelli. Cuore e circolaz., 8 : 362— 368. Pfister A. 1937. Beobachtungen an eineiigen Zwillingspaaren. Arch. J. Klaus-Stift., 12 : 587—629. Planansky K. a. G. Allen. 1953. Heredity in relation to variable resistance to pulmonary tuberculosis. Amer. J. Hum. Gen., 5 : 322— 349. Platt R. a. R. Lawton. 1956. The retinae of monovular and binovular twins. Ann. Hum. Gen., 21 : 132—134. Pоlitzer G. 1955. Theorie der Doppelmissbildungen. Ztschr. mikrosk. anat. Forsch., 61 : 590—623. Poll H. 1914. Uber Zwillingsforschung als Hilfsmittel menschlicher Erbkunde. Ztschr. f. Ethnol., 46 : 87—108. Poll H. 1930. Zwillinge in Dichtung und Wirklichkeit. Ztschr. ges. Neurol. u. Psychiatr., 128 : 423—474. Potter E. 1953. Multiple pregnancies and cone joined twins. In: Pathology of the foetus and the newborn. Chicago : 172—198. Prell H. 1927. Uber doppelte Brunstzeit und verlangerte Tragzeit bei den einheimlichen Arten der Mardergattung Martes Pinal. Zool. Anz., 74 : 122—128. Presslek K. 1911. Beobachtungen und Versuche uber den normalen und inversen Situs viscerum et cordis bei Anurenlarven. Arch. f. Entw.-Mech., 32 : 1—35. Price B. 1950. Primary bases in twin studies. A review of prenatal and natal difference-producing factors in monozygotic pairs. Amer. J. Hum. Gen., 2 : 293—352. Pryor J. W. 1936. Ossification as additional evidence in differentiating identicals and fraternals in multiple birth. Amer. J. Anat., 59 : 409—423. Pryor J. W. 1939. Normal variation in the ossification of bones. J. Hered., 30 : 249—255. Pryor J. W. 1948. The Badgett quadruplets. J. Hered., 39 : 3. Quelprud Th. 1932. Untersuchungen der Ohrmuschel von Zwillingen. Ztschr. ind. Abstamm.- u. Vererbungsl., 62 : 160—165. Quelprud Th. 1940. Ear pit and its inheritance. J. Hered., 31 : 379—384. Quick A. a. J. Соnway. 1949. Hemophilia in twins. Amer. J. Med., 7 : 841—843. Randall T. E. a. С. M. Rick. 1945. A cytogenetic study of polyembryony in Asparagus officinalis L. Amer. J. Bot., 32 : 560. Raneу E. T. 1938. Reversed lateral dominance in identical twins. J. Exper. Psychol., 23 : 304—311. Raneу E. T. 1939. Brain potentials and lateral dominance in identical twins. J. Exper. Psychol., 24 : 21—39. Raubitschek H. 1955. Das Geschick identischer Zwillingspaare. Wien. klin. Wochenschr., 67 : 74—76. Rawlings E. a. R. Warwick. 1951. Conjoined twins. J. Obst. Gynaecol., Brit. Emp., 58 : 452—455. Record R. a. Th. McKeown. 1951. Congenital malformations of the central nervous system. Ann. Eugen. 15 : 285—292. Reed Sh., 1938. Uniovular twins in mice. Science, 13. Reinhardt. 1912. Ein Fall von Situs viscerum inversus totalis bei Zwillingen (Rekruten). Dtsch. Militararztl. Ztschr., 41 : 931—934. Reitman H., E. Smith a. J. Geller. 1953. Separation and survival of xiphopagus twins. J. Amer. Med. Ass., 153, 15 : 1360—1362. Rempel J. C. 1954. Siamese twin embryos of the black widow spider, Latrodectus magtaus (Fabr.). Canad. J. Zool., 32 : 240—243. Rhodes A. 1943. Pseudohermaphroditism in twins; report of case. Arch. Pediatr., 60 : 529—536. Richter J. 1926. Zwillings- und Mohrlingsgeburten bei unseren Landwirtschaftlichen Haussaugetieren. Ztschr. Zucht., 29. Riddle 0. 1923. On the cause of twinning and abnormal development in birds. Amer. J. Anat., 32 : 199—252. Rife D. 1933a. Genetic studies of monozygotic twins. I. A diagnostic formula. J. Hered., 24 : 339—345. Rife D. 1933b. Genetic studies of monozygotic twins. II. Finger patterns and eye-color as criteria of monozygosity. J. Hered., 24 : 406—414. Rife D. 1939. A study of twins. Sci. Monthly, 48 : 236—245. Rife D. 1940. Handedness, with special reference to twins. Genetics, 25 : 178—186. Rife D. 1943. Handedness and dermatoglyphics in twins. Hum. Biol., 15 : 46—54. Rife D. 1952a. Twins and research. Acta Gen. Med. Gemellolog. 1 : 14—24. Rife D. 1952b. A quantitative analysis of finger prints, Hum. Biol., 24 : 1. Rife D. a. H. Cummins. 1943. Dermatoglyphics in twins. Hum. Biol., 15, 1. Rife D. a. M. Hunstead. 1946. A pair of albini twins. J. Hered., 37 : 3—4. Robertson Al. 1950. Some observations on experiments with identical twins in dairy cattle. J. Genet., 50 : 32—35. Roeder R. 1950. Zwillingsforschung und das Erblichkeits-Umweltsproblem. Dtsch. Gesundheitsw., 5 : 342—345. Rohlfs D. 1928. Die Eiigkeitsdiagnose aus der Ahnlichkeit bei neugeborenen Zwillingen. Arch. Gynak., 133 : 841—848. Roman-Goldzieher R. R. 1945. Untersuchungen der Schrift und des Schreibens von 283 Zwillingspaaren. Schweiz. Ztschr. Psychol. Anwend. Beih., 6 : 29—55. Rossle R. 1937. Zur Frage der Ahnlickeit des Windungsbildes an Gehirnen von Blutsverwandten, besonders von Zwillingen. Sitzungsber. Preuss. Akad. Wiss., Phys.-Math. Kl., 13—14 : 146—168. Rossle R. 1939. Erbpathologie des Menschen. C. R. Congr. Intern. Pathol. Comp., Rome, 1. Rossle R. 1940. Die pathologische Anatomie der Familie. Berlin. Rossle R. 1950. Weitere Beitrage zur anatomischer Ahnlichke Blutsverwandter. Dtsch. Gesundheitsw., 5 : 1442—1444. Rоmanus T. 1947. Psoriasis in twins. Hereditas, 33 : 297—300. Rоsanоff A. 1934. The etiology of so-called schizophrenia psychoses Amer. J. Psychiatr., 91 : 247—286. Rоsanоff A. a. Inman-Kane. 1934. Relation of premature birth and underweight condition at birth to mental deficiency. Amer. J. Psychiatr., 13 : 829. Ruhl H. 1938. Uber Polydaktylie bei Zwillingen. Zbl. Gynak., 62 : 2706—2709. Rumpe K. 1891. Uber einige Unterschiede zwischen eineiigen und zweieiigen Zwillingen. Ztschr. Geburtsh. u. Gynak., 22. Saudeк R. u. E. Seaman. 1933. Die Handschrift eineiiger, getrennt erzogener Zwillinge. Character, 2 : 66—81. Schachter M. et H. Chatenet. 1953. Nouvelle contribution a l’etude du test de Rorschach chez des jumeaux. Acta Gen. Med. Gemellolog., 2 : 69—86. Schatz F. 1882. Eine besondere Art von oinseitiger Polyhydramnie mil anderseitiger Oligohydramnie bei eineiigen Zwillingen. Arch. Gynak. (Berlin), 19 : 329—369. Schatz F. 1884—1910. Die Gefassverbindungen der Plazentarkreislaufe eineiiger Zwillinge, ihre Entwicklung und ihre Folgen. Arch. Gynak., 24, 26, 29, 30, 53, 55, 58, 60, 92. Schermer S. 1936. Eineiige Zwillinge beim Pferd. Zuchtungskunde, 11 : 395—401. Schiff F. u. O. Verschuer. 1933. Serologische Untersuchungen an Zwillingen. Ztschr. Morph, u. Anthrop., 32 : 244—249. Schiller M. 1937. Zwillingsprobleme, dargestellt auf Grund von Untersuchungen an stuttgarter Zwillingen. Ztschr. menschl. Vererbungsl., 20 : 284—337. Schiller M. 1942. Realitat und Problematik der menschlichen Handfurchen. . . Ztschr. menschl. Vererb.- u. Konstitutionsl., 52 : 129 сл. Schinz H. R. 1945. Konkordanz, Diskordanz und Penetranz bei eineiigen Zwillingen. Arch. J. Klaus.-Stift., 20: 13—25. Schinz H., U. Сосchi, J. Neuhaus. 1948. Die Vererbung des Krebses beim Menschen. Arch. J. Klaus-Stift., 23. Schlaginhaufen O. 1940. Die Vierlingsgeschwister Gehri und ihr Verwandtschaftskreis. Eine familienanthropologische Untersuchung. Arch. J. Klaus-Stift., 15 : 309—398. Schieip W. 1929. Die Determination in der Primitiventwicklung. Leipzig. Schmidt O., 11. Manz u. Traenckner. 1951. Serologische Untersuchungen bei Zwillingen usw. Ztschr. Ges. gericht. Med., 40 (3) : 197—223. Schultz A. H. 1956. The occurrence and frequency of pathological and teratological conditions of twinning among non-human primates. In: Primatologia. Handbuch der Primatenkunde. Basel— N. Y. : 1005—1007. SchuItze O. 1894. Die kunstliche Erzeugung von Doppelbildungen bei Froschlarven, mit Hilfe abnormer Gravitationswirkung. Arch. Entw.-Mech., 1 : 269—305. Schwaegerle F. 1938. Irisstructur und Augenfarbe bei ein- und zweieiigen Zwillingen. Ztschr. menschl. Vererb.- u. Konstitutionsl., 22 : 545—577. Schwalbe E. 1907. Die Morphologic der Missbildungen des Menschen und der Tiere. Teil II. Die Doppelbildungen. Jena : 410 Schwalbe E. 1923. Missbildungen. In: Pathologische Anatomic Her. L. Aschoff. Jena, I : 324—357. Schwarzburg W. 1927. Statistische Untersuchungen uber den menschlichen Scheitelwirbel und seine Vererbung. Ztschr. Morph, u. Anthrop., 26. Schwesinger G. 1940. Five little Dionnes and how they grew J. Hered., 31 : 145—150. Schwesinger G. 1952. The effect of differential parent-child relation on identical twins resemblance in personality. Acta Gen. Med. Gemellolog., 1 : 40—47. Schwesinger G. 1956. Heredity and environment in the development of intelligence. Novant’anni delle leggi mendeliane, Roma : 199—205. Schwind J. L. 1942. Spontaneous twinning in the amphibia. Amer. J. Anat., 71 : 117—151. Seemann M. 1937. Die Bedeutung dor Zwillingspathologie fur die Erforschung von Sprachleiden. Arch. Sprach.- u. Stimmenheilk., 1 : 88—98. Seidel Fr. 1952. Die Entwicklungspotenzen einer isolierten Blastomere des Zweizellenstadiums im Saugetierei. Naturwissenschaften, 39 : 355—356. Siebenthal R. 1945. Les accouchements gemellaires a la maternite de Geneve do 1934 a 1943. Monatschr. Geburtshilfe u. Gynak., 120 : 225—332. Siebert E. O. 1937. Anatomische Untersuchungen uber die Ahnlichkeit bei eineiigen und zweieiigen Zwillingen. Ztschr. Anat. u. Entwicklungsgesch., 108 : 1—30. Sieder H. 1938. Uber die Augenlider bei Zwillingen. Ztschr. menschl. Vererb.- u. Konstitutiosl., 22 : 304—341. Siemens H. 1924. Die Zwillingspathologie. Ihre Bodeutung, ihre Methodik, ihre bisherige Ergebnisse. Berlin. Siemens H. 1927. Diagnosis of identity in twins. J. Hered., 18 : 201 — 209. Siemens H. 1952. Zur Geschichte der Zwillingsmethode. Ztschr. menschl. Vererb.- u. Konstitutionsl., 31 : 171 —173. Silоw R. a. S. Stephens. 1944. «Twinning» in cotton. J. Hered., 35 : 76—78. Silvestri F. 1906. Contribuzioni alla connoscenza biologica degli Imenotteri Parassiti. I. Biologia del Litomastrix truncatellus. Ann. R. Scuola Agric. portici, 6 : 1—51. Simonds B. 1957a. Twin research in tuberculosis. Eugen. Rew., 49 : 25—32. Simonds B. 1957b. The collection of 300 twin index cases for a study of teberculosis in twins and their families. Acta genet. et statist. med., 7 : 42—52. Slater E. 1953. Psychotic and neurotic illness in twins. London. Smith C. 1949. Psychological studies in twin differences. Lund : 1 —251. Smith S. a. L. Penrose. 1955. Monozygotic and dizygotic twin-diagnosis. Ann. Hum. Gen., 19 : 273—289. Sommering T. S. 1791. Abbildung und Beschreibung einiger Missgeburten. Mainz. Sоntag L. a. E. Reinоlds. 1944. Ossification sequences in identical triplets. J. Hered., 35 : 57—64. Spemann H. 1901. Entwicklungsphysiologische Studien am Tritonei. I. Arch. Entw.-Mech., 12 : 224—264. Spemann H. 1902. Entwicklungspliysiologische Studien am Tritonei. II. Arch. Entw.-Mech., 15 : 448—534. Spemann H. 1903. Entwicklungsphysiologische Studien am Tritonei. III. Arch. Entw.-Mech., 16 : 551—631. Spemann H. 1936. Experimentelle Bcitrage zu einer Theorie der Entwicklung. Springer-Verlag. Spemann H. u. H. Falkenberg. 1919. Uber asymmetrische Entwicklung und Situs inversus viscerum bei Zwillingen und Doppel-bildungen. Arch. Entw.-Mech., 45 : 371—422. Splinder P. 1955. Ausdruck und Verhalten erwachsener Zwillinge. Acta Gen. Med. Gemellolog., 4 : 32—61. Stadler H. 1955. Disparity in the cardiac status of monozigotic twins. J. Pediatr. 47 : 353—356. Steiner Fr. 1935. Nachgeburtsbefunde bei Mehrlingen und Ahnlichkeitsdiagnose. Arch. Gynak., 159 : 509—523. Steinmann P. 1943. Uber seitenverkehrte Zwilling und uber die Symmetrie von Doppel-Monstrositaten. Arch. J. Klaus-Stift., 18 : 703—707. Stewens W. 1937. A test for uniovular twins in mice. Ann. Eugen., 8 : 70—73. Stockard С. R. 1921. Developmental rate and structural expression: an experimental study of twins, «double monsters» and single deformities and the interaction among embryonic organs during their origin and development. Amer. J. Anat., 28 : 115—277. Stocks P. 1930. A biometric investigation of twins and their brothers and sisters. Part I. Ann. Eugen., IV : 49—108. Stocks P. 1933. A biometric investigation of twins and their brothers and sisters. Part. II. Ann. Eugen., V : 1—55. Stocks P. 1952. Recent statistics of multiple births in England and Wales. Acta Gen. Med. Gemellolog., 1 : 8—13. Stocks P. 1953. Multiple birth frequency according to parity and maternal age. Acta Gen. Med. Gemellolog., 2 : 113—117. Stormont C. 1954. Research with cattle twins. In: Kempthone O. et al. Statistics and mathematics in biology. Ames, Jowa State College Press : 407—418. Stormont C., W. Weir a. L. Lane. 1953. Erythrocyte mosaicism in a pair of sheep twins. Science, 118, 3075 : 695—696. Strandskov tH. a. G. Diederch. 1945. The Rh blood factor among twins. Hum. Biol., 17 : 195—206. Strandskоv H. a. E. Edelen. 1946. Monozygotic and dizygotic twin birth frequencies in the total the «white» and «colored» U. S. populations. Hum. Gen., 31 : 438—446. Strandskоv H. a. G. Siemens 1946. An analysis of the sex rations among single and plural births in the total, the «white» and the «colored» U. S. populations. Amer. J. Phys. Anthrop., N. S., 4 : 491—501. Strandskоv H. a. D. Оndina. 1947. A comparison of the percentages of still-births among single and plural births in the total etc. Amer. J. Phys. Anthrop., N. S., 5 : 40—54. Strasburger. 1873. Uber Polyembryonie. Jen. Ztschr. Naturw., 12. Strayer L. C. 1930. Language and growth: the relative efficacy of early and deferred vocabulary training, studies, by the method of co-twin control. Gen. Psychol. Monogr., 8 : 209—219. Streeter G. L. 1919. Formation of single-ovum twins. J. Hopkins Hosp. Bull., 30 : 235—238. Streeter G. L. 1924. Single-ovum twins in the pig. Amer. J. Anat., 34 : 183—194. Sturkie P. 1946. The production of twins in Gallus domesticus. J. Exp. Zool., 101 : 1. Swelt W. et al. 1940. Early recognition of the free-martin condition in heifers twinborn with bulls. J. Agric. Res., 61 : 587—624. Tannreuter G. W. 1919. Partial and complete duplicity in chick embryos. Anat. Rec., 16 : 355—367. Thomes N. 1921. Twins in the Yoruba country. Man, 21. Thоrndike E. L. 1905. Measurements of twins. Arch, phylosoph. psychol. a. Sci. met., 1 : 1—64. Tisserand-Perrier M. et Вlaizоt. 1955. Le dessin, mode d’investigation de la personnalite chez les jumeaux identiques. Acta Gen. Med. Gemellolog., 4 : 261—274. Tоmilin M. J. a. R. M. Jerkes 1935. Chimpanzee twins: Behavioral relations and development. J. Gen. Psychol., 46 : 239— 263. Torgersen J. 1948. Concordant situs inversus in dizygotic twins. J. Hered., 39 : 293—294. Torgersen J. 1950. Situs inversus, asymmetry and twinning. Amer. J. Hum. Gen., 2 : 361—370. Turner C. L. 1956. Twinning and reproduction of twins in Pelmatohydra oligactis. Biol. Bull., 111 : 269—277. Turpin R. 1942. Etude correlative des sinus frontaux des jumeaux. C. R. Soc. Biol., 136 : 203—205. Turpin R. (Red.). 1955. La progenese. Facteurs preconceptionnels du developpement de l’enfant. Paris: 1—703. Turpin R. et J. Pittоn. 1938. Gemellite en miroir et double noyau d’ossification du semilunaire. Bull. Soc. Pediatr. Paris, 6 : 390—395. Turpin R. et M. Garatzali. 1941. Recherche sur les correlations leucocytaire des jumeaux. Proc. Gen. Congr. Edinburgh, 7 : 301. Turpin R. et M. Zazzo. 1943. Elude psychologique et motrice de jumeaux monozygotiques en miroir. Bull. Acad. Med., 127, 21/22 : 344—346. Turpin R. et G. Вernуer. 1947. Cancer et gemellite. Commun. au XI congres des pediatres de langue francaise. Lyon, 2-e vol. : 173. Turpin R. el M. Schutzenberger. 1950. Sur la masculinite a la naissance dans les grosses multiples. C. R. Acad. Sci. Paris. 231, 20 : 1098—1099. Turpin R. el M. Schutzenberger. 1952a. Sexe et gemellite. Semaine des Hopitaux, 28, 44 : 1844—1848. Turpin R. el M. Schutzenberger. 1952b. Progenese et gemellite. Acta Gen. Med. Gemellolog., 1 : 317. Twisselmann F. 1939. Experiences de scission precoce de l’aire einbryonnaire chez le poulet. Arch. Biol., 49. Uehlinger E. u. Кunsch. 1938. Uber Zwillingstuberkulose. Beitr. klin. Tuberk., 92 : 275. Uppenborn W. 1933. Untersuchungen uher die Trachtigkeit der Stuten, mit einem Anhang uber Zwillingsgeburten beim Pferd. Ztschr. Zucht., 28 : 1—27. Valentin G. 1851. Ein Beitrag zur Entwicklungsgeschichte der Doppelmissbildungen. Vierordts Arch, physiol. Heilk., 10 : 1—40. Vejdowsky Fr. 1892. Entwicklungsgeschichtliche Untersuchungen. Prag. Verschuer O. 1927. Studien an 102 eineiigen und 45 gleichgeschlechtlichen zweieiigen Zwillings- und an 2 Drillingspaaren. Ergebn. d. inn Med. u. Kinderheilk., 31 : 35 120. Verschuer O. 1929. Zur Frage der Asymmetrie des menschlichen Korpers. Ztschr. Morph. u. Anthrop., 27 : 171—178. Verschuer O. 1931/32. Ergebnisse der Zwillingsforschung. Verhandl. Ges. phys. Anthrop., VI : 1—65. Verschuer O. 1932. Die biologischen Grundlagen der menschlichen Mehrlingsforschung. Ztschr. ind. Abstamm.- u. Vererbungsl., 61 : 147—205. Verschuer O. 1934a. Zur Erbbiologie der Fingerleisten, zugleich ein Beitrag zur Zwillingsforschung. Ztschr. ind. Abstamm.- u. Vererbungsl., 67 : 299—301. Verschuer O. 1934b. Die Erbbedingtheit des Korperwachstums. Zeitschr. Morph. u. Anthrop., 34 : 398—412. Verschuer O. 1934c. Methoden der Erbforschung beim Menschen. Naturwissenschaften, 22 : 765—771. Verschuer O. 1937. Erbpathologie. Ein Lehrbuch fur Arzte und. Medizinstudierende. Leipzig. Verschuer O. 1939. Twin research from the time of F. Galton to the present day. Proc. Roy. Soc., Ser. B., 128, 850 : 62—81. Verschuer O. 1952. Erbforschung am Menschen — neue Ergebnises und Erkenntnisse. Dtsch. Med. Wochenschr., 77, 41 : 1245— 1248. Verschuer O. 1954. Wirksame Faktoren im Leben des Menschen. Beobachtungen an ein- und zweieiigen Zwillingen durch 25 Jahre. Wiesbaden: 1—288+105. Verschuer O. 1955. Tuberkulose Zwillinge. Nachuntersuchung nach 20 Jahren. Dtsch. Med. Wochenschr., 80, 45 : 1635—1637. Verschuer O. 1956. Die Bedeutung der erblichen Veranlagung fur den Ablauf der Tuberkulose. Nachuntersuchung nach 20 Jahren an einer Serie von tuberkulosen Zwillingen. Novant’ anni delle leggi mendeliane, Roma. Verschuer 0., W. Kinkelin u. V. Zipperlen. 1927. Die vererbungsbiologische Zwillingsforschung. Ergebn. inn. Med. u. Kinderheilk., 31 : 35—120. Verschuer О. u. E. Коber. 1940. Dio Frage der erblichen Disposition zum Krebs. Ztschr. f. Krebsforsch., 50 : 5—14. Verschuer О. u. V. Zipperlen 1929. Die erb- und umweltbedingte Variabilitat der Herzform. (Nach Rontgenaufnahmen des Herzens bei Zwillingen). Ztschr. klin. Med., 112 : 69—92. Virchow R. 1891. Xiphodymen Gebruder Tocci. Ztschr. Ethnol., 23 : 245—246. Vogel F. 1956. Dermathologische Beobachtungen an eineiigen Zwillingen: Vitiligo, Ichthyosis, Psoriasis. Ztschr. Haut. u. Ge-schlechtskr., 20, 1 : 1—4. Vogt A., H. Wagner, H. Richner, G. Мeуer. 1939. Das Senium bei eineiigen und zweieiigen Zwillingen. Die Erbentstehung bisher exogen und durch «Abnutzung» erklarten Altersleiden. Arch. J. Klaus-Stift., 14 : 475—597. Waardenburg P. J. 1932. Das monschliche Auge und seine Erbanlagen. Haag : 557—576. Waardenburg P. J. 1950 Twin research in ophthalmology. Documenta Ophthalm., 4 : 154—199. Waardenburg P. J. 1955. Ein autosomal-dominantes Strukturmerkmal der menschlichen Iris. Acta Gen. Med. Gemellolog., 4 : 161- 179. Waddington С. H. 1941. Twinning in chick embryos. J. Hered., 32 : 268—270. Wagenseil F. 1931. Zwei Mitteilungen uber die erbbiologische Bedeutung der eineiigen Mehrlinge. Ztschr. Konstitutionsl., 15 : 632— 645. Walsh R. a. 0. Kooptzoff. 1955. A study of twins blood groups and other data. Austral. J. Exper. Biol. Med. Sci., 33 : 189—198. Wardlaw G. 1955. Embryogenesis in plants. : 292—293. Warkanу J. a. Selkirk. 1955. Discordant monozygotic twins. Amer. J. Dis. Child., 89 : 144—148. Waterhouse J. 1953. Twin studies. In: Clinical genetics, ed. by A. Sorsby : 35—46. Weber J. M. 1940. Polyembryony. Bot. Rev., VI, 11. Weber W. 1945. Untersuchungen uber die Haufigkeit und Erblichkeit der Disposition zu Zwillingsgeburten beim Simmentalerrind. Arch. J. Klaus-Stift., 20 : 3—4. Wehefritz E. 1925. Uber die Vererbung der Zwillingsschwangerschaft. Ztschr. Konstitutionsl., 11 : 554—575. Weinberg W. 1903. Methode und Ergebnis der Erforschung der Ursachen der Mehrlingsgeburten. Virch. Arch. : 171. Weinberg W. 1928. Zur Berechnung der Haufigkeit eineiiger Zwillinge. Arch. Gynank., 133 : 289—290. Weinberg W. 1934. Differenzmethode und Geburtenfolge bei Zwillingen. Genetica, 16 : 382—388. Weitz W. 1936. Die Vererbung innerer Krankheiten. Stuttgart. Wells L. J. 1945. A case of iliothoracopagus (dicephalus tribrachiustripus) with a consideration of the «budding» ana «fission» theories of twinning. Anat. Rec., 93 : 1—22. Wendt G. 1955. Der individuelle Musterwert der Fingerleisten und seine Vererbung. Acta Gen. Med. Gemellolog., 4 : 330—337. Wenner R. 1947. Uber den plazentaren Blutkreislauf bei eineiigen Zwillingen. Schweitz. med. Wochenschr., 4 : 140—141. Wenner R. 1952. Les jumeaux homozygotes univitellins. Rev. franc. gynec. et obst., 5—6 : 174—184. Werner M. 1935. Blutzuckerregulation und Erbanlage. Dtsch. Arch, klin. Med., 178 : 308—338. Werner M. 1938. Die Erb- und Umweltbedingtheit der Unterschiede bei der vitalen Lungenkapazitat usw. Ztschr. menschl. Vererb.-u. Konstitutionsl., 24 : 293—305. Wessel E. 1926. Experimentell erzeugte Duplicitas cruciata bei Triton. Roux’Arch., 107 : 480—556. Wetner M. 1937. Erb- und Umweltunterschiede in der Vitalkapazitat der Lungen. Untersuchungen an 70 Zwillingspaaren. Ztschr. ind. Abstamm.- u. Vererbungsl., 73 : 527—531. Wetzel G. 1900. Drei abnorm gebildete Eier von Tropidonotus natrix Anat. Anz., 18 : 425—440. Wiese N. et al. 1939. An electrocardiographic study of twins. Amer Heart Journ., 17 : 701. Wilder H. 1904. Dublicate twins and double monsters. Amer. J. Anat., 3 : 387. Wilson J. a. D. Gregory. 1931. Sextuplet lambs. J. Hered., 22 : 229—230. Wislосki G. B. 1939. Observations on twinning in marmosets. Amer. J. Anat., 64 : 445. Witschi E. 1934. Appearance of accessory «organizers» in overripe eggs of the frog. Proc. Soc. Exp. Biol. Med., 31 : 419—420. Wolff E. 1948. La science des monstres. Paris : 1—265. Woellwarth C. 1950. Experimentelle Untersuchungen uber den Situs inversus der Eingoweide und der Habenula des Zwischenhirns bei Amphibien. Roux’Arch., 144 : 178—256. Wricke G. 1954. Die Manifestierung der erblich bedingten Polyembryonie beim Lein in ihrer Abhangigkeit von Aussenfaktoren. Biol. Zbl., 75 : 49—88. Yerkes R. M. 1934. Multiplebirths in anthropoid apes. Science, 79 : 430—431. Yerushalmy J. a. S. Sheear. 1940a. Studies on twins. I. The relation of order of birth and age of parents to the frequency of like-sexed and unlike-sexed twin deliveries. Hum. Biol., 12 : 95—113. Yerushalmy J. a. S. Sheear. 1940b. Studies on twins. II. On the early mortality of like-sexed and unlike-sexed twins. Hum. Biol., 12 : 247—263. Zarapkin S. R. 1939. Autonome Variation der Merkmale des Individuums und Analyze der Divergenzerscheinungen bei Zwillingen. Ztschr. Konstitutionsl., 23 : 636—657. Zazzо R. 1955. Sur le postulat de la comparabilite duns la methode des jumeaux. Acta Gen. Med. Gemellolog., 4 : 180—189. Zоja R. 1895. Sullo sviluppo dei blastomeri isolati dalle uova di alcune medusae. Arch. Entw.-Mech., 1 : 578—618.
Последние комментарии
4 часов 18 минут назад
10 часов 2 минут назад
11 часов 9 минут назад
12 часов 7 минут назад
12 часов 21 минут назад
21 часов 31 минут назад